








 114 / 2894
114 / 2894

muscular. Las moléculas de transmisor que no reaccionan inme-
diatamente con un receptor o las que son liberadas después de la
unión al receptor son destruidas casi instantáneamente por la
acetilcolinesterasa en la hendidura sináptica. La acetilcolinesterasa
de la unión es la forma proteica A12 o asimétrica, sintetizada en el
músculo bajo la placa terminal. La acetilcolinesterasa (clasificación
de enzimas 3.1.1.7) es el tipo B de enzima carboxilesterasa. Hay
una concentración menor de la misma en la zona externa a la unión
sináptica. La enzima se excreta desde el músculo pero permanece
adherida al mismo mediante filamentos finos de colágeno que
están enganchados a la membrana basal
13,17
. La mayoría de las
moléculas de acetilcolina liberadas desde el nervio pasan inicial-
mente entre las enzimas para alcanzar los receptores postsinápti-
cos, aunque conforme son liberadas del receptor, se topan
invariablemente con la acetilcolinesterasa y son destruidas. En
circunstancias normales, una molécula de acetilcolina reacciona
sólo con un receptor antes de ser hidrolizada. La acetilcolina es un
potente mensajero, pero su acción dura muy poco tiempo porque
es destruida en menos de 1 milisegundo después de ser liberada.
Existen enfermedades congénitas y adquiridas que están
relacionadas con una actividad alterada de la enzima acetilcolines-
terasa. La ausencia congénita de la enzima excretada (en ratones
knock-out
, es decir, defectivos para un gen) conlleva una alteración
en el mantenimiento del sistema neuronal motor y en la organiza-
ción de las ramas nerviosas terminales
51
. Hay muchos síndromes
que se deben a anomalías congénitas de la función de la colineste-
rasa y que provocan enfermedades neuromusculares cuyos signos
y síntomas habitualmente se asemejan a los de la miastenia grave
o síndromes miasténicos
52,53
. La denervación disminuye la acetil-
colinesterasa en las zonas sináptica y extrasináptica
31
. Otras enfer-
medades adquiridas que afectan a las colinesterasas tienen que ver
con la inhibición crónica de la acetilcolinesterasa por pesticidas
organofosforados o gases nerviosos (p. ej., sarín) o con el trata-
miento crónico con piridostigmina administrado como profilaxis
contra el envenenamiento por gas nervioso
54,55
. Los síntomas, que
van desde la fatiga crónica hasta la debilidad muscular, se han
atribuido a la inhibición crónica de la colinesterasa, por tanto se
ha infravalorado la importancia de la acetilcolinesterasa en la
función neuromuscular normal y anómala.
Receptores postsinápticos de acetilcolina
La similitud entre los RACh de bastantes especies y la abundancia
de receptores de acetilcolina en el pez eléctrico
Torpedo
han faci-
litado mucho la investigación en esta área. La disponibilidad del
ácido ribonucleico mensajero (ARNm) de seres humanos y de
otras especies, así como la del ácido desoxirribonucleico (ADN) ha
posibilitado el estudio del receptor en sistemas artificiales como los
ovocitos de ranas y en células de mamíferos que no expresan el
receptor, como COS o fibroblastos. También es posible, utilizando
técnicas moleculares, producir mutaciones en los receptores para
simular situaciones patológicas y estudiar la función del receptor
en estos sistemas artificiales. Con el uso de estas técnicas y de otras
relacionadas se ha avanzado en el conocimiento de la síntesis,
composición, función biológica y mecanismos que subyacen en las
respuestas fisiológicas y farmacológicas de los RACh
56-59
. Existen
tres isoformas de receptores postsinápticos: una forma sináptica o
madura, un receptor extrasináptico o inmaduro (fetal), y el recep-
tor neuronal
a
7 descrito más recientemente (v. «Biología de los
receptores nicotínicos de acetilcolina pre y postsinápticos»)
1-3
. Sin
embargo, las diferencias entre los subtipos de receptores pueden
obviarse en una explicación general sobre la función de los recep-
tores en la transmisión neuromuscular.
Los RACh son sintetizados en las células musculares y se
almacenan en la membrana de la placa terminal mediante una
proteína especial de 43 kd conocida como rapsina. Esta proteína
citoplásmica se asocia al receptor de acetilcolina en una proporción
de 1:1
31
. Los receptores, formados por cinco subunidades proteicas,
están dispuestos como las cuadernas de un barril dentro de un
receptor cilíndrico con un poro central para el canal iónico. En la
figura 4-4 se puede ver un esquema de los factores fundamentales.
114
Fisiología y anestesia
I
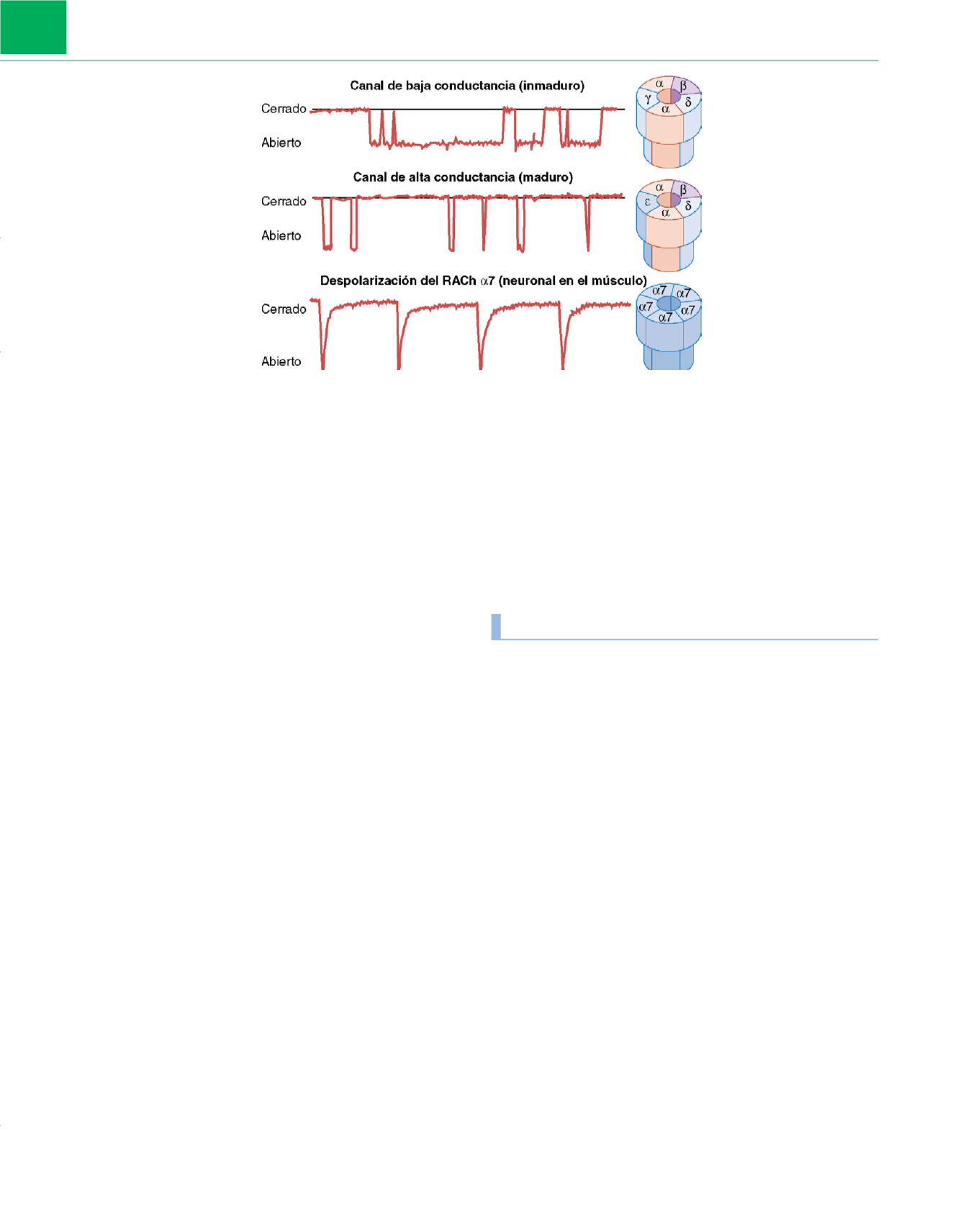
Figura 4-4
Esquema de los canales de los receptores del RACh
(derecha)
y de los trazados de registros de muestras celulares de la apertura de los canales de los
receptores
(izquierda)
. El receptor maduro, o de la sinapsis, está formado por dos subunidades
a
y por una subunidad
b
,
d
, y
ε
, respectivamente. La forma
inmadura, fetal o extrasináptica, está constituida por dos
a
y por una subunidad
b
,
d
y
g
, respectivamente. Por tanto, el último se denomina receptor de subunidad
g
.
Recientemente se ha descrito en el músculo un receptor neuronal que consta de cinco subunidades
a
7. Estas subunidades están dispuestas alrededor del canal
catiónico central. La isoforma inmadura que contiene la subunidad
g
muestra tiempos de apertura prolongados y corrientes del canal de amplitud baja. La isoforma
madura que contiene la subunidad
ε
muestra tiempos de apertura más cortos y unas corrientes del canal elevadas durante la despolarización. La sustitución de la
subunidad
ε
por la subunidad
g
produce el tipo de canal de apertura rápida y conductancia elevada. Como era de esperar, la aplicación de acetilcolina al RACh
a
7
también provoca una corriente rápida interior con un descenso acelerado. Todos estos acontecimientos despolarizantes son insensibles al tratamiento con atropina,
aunque son sensibles al tratamiento con
a
-bungarotoxina o con relajantes musculares, los cuales bloquean el flujo de corriente.