








 115 / 2894
115 / 2894

La proteína receptora tiene un peso molecular de unos 250.000 dal-
tons. Cada receptor tiene cinco subunidades. El receptor maduro
consta de las subunidades
a
,
b
,
d
y
ε
, y el receptor fetal (inmaduro
y extrasináptico) de las
a
,
b
,
d
y
g
; existen dos subunidades
a
y una
subunidad de cada una de las otras. El RACh neuronal
a
7 consta
de cinco subunidades
a
7
2
. Cada una de las subunidades de todos
los receptores está compuesta aproximadamente por 400 o 500 ami-
noácidos. El complejo proteico del receptor atraviesa por com-
pleto la membrana y protruye más allá de la superficie extracelular
de la misma y en el interior del citoplasma. El sitio de unión para
la acetilcolina está en cada una de las subunidades
a
, y constituye
la zona competitiva entre los agonistas y los antagonistas del recep-
tor. Los agonistas y antagonistas son atraídos hacia el sitio de unión
y pueden ocupar el lugar, que se localiza cerca del residuo de cis-
teína (exclusivo de la cadena
a
) en la posición 192-193 de la cadena
de aminoácidos de la subunidad
a
60
. La
a
-bungarotoxina radio-
marcada procedente de la cobra, que se utiliza para cuantificar el
receptor, se une a la región heptapeptídica 189-199 de la subuni-
dad
a
61
. La neurregulina-1
b
(NR
b
-1) derivada de la motoneurona,
descrita inicialmente con actividad inductora del receptor de acetil-
colina (AIRA), induce la transcripción del gen del RACh en los
mionúcleos subsinápticos mediante la activación de receptores
ErbB
62
.
Síntesis y estabilización de los receptores
postsinápticos
El tejido muscular está formado a partir del mesodermo, y al prin-
cipio aparece como mioblastos. Los mioblastos se fusionan para
producir miotubos, los cuales tienen, por tanto, múltiples núcleos.
Conforme los miotubos van madurando, se desarrolla el sarcómero,
que es el elemento contráctil del músculo, compuesto por actina y
miosina
63
. La proteína
b
-integrina parece esencial para la fusión de
mioblastos y para el ensamblaje del sarcómero
64
. Poco tiempo
después, los axones de los nervios motores crecen en el interior del
músculo en desarrollo, y estos axones aportan señales derivadas del
nervio (es decir, factores de crecimiento), incluidas agrina y
neurregulinas (NR
b
-1 y NR
b
-2), que son fundamentales para la
maduración de los miotubos hacia el músculo
62
. La agrina es una
proteína del nervio que estimula la diferenciación postsináptica
mediante la activación de una cinasa muscular específica (MuSK),
una tirosín-cinasa expresada de forma selectiva en el músculo. Con
la señalización por la agrina, los receptores de acetilcolina, que han
estado dispersos por toda la membrana muscular, se agrupan en el
área inmediatamente por debajo del nervio. La agrina, junto a otros
factores de crecimiento (neurregulinas, etc.), induce también el
agrupamiento de otras proteínas críticas derivadas del músculo,
incluidas MuSK, rapsina y proteínas ErbB, todas ellas necesarias
para la maduración y estabilización de los receptores de acetilcolina
en la unión (fig. 4-5). Además de los efectos sobre la diferenciación
postsináptica, la agrina y la MuSK también exhiben efectos en la
diferenciación presináptica. La agrina y MuSK inducen señales
retrógradas que instruyen a los axones para provocar el crecimiento
neuronal y la diferenciación terminal
19
. Sin embargo, a día de hoy
se conoce menos el desarrollo presináptico de la unión neuromus-
cular que el desarrollo postsináptico. Justo antes y un poco después
del nacimiento, los RACh que contienen la subunidad
g
inmadura
son convertidos en los receptores maduros que contienen la subu-
nidad
ε
. Aunque no se conoce con exactitud el mecanismo de este
cambio,parece que está involucrada la neurregulinaNR
b
-1,también
llamada AIRA, que se une a uno de los receptores ErbB
44,62-65
.
Electrofisiología básica
de la neurotransmisión
El conocimiento de las técnicas de electrofisiología ha progresado al
mismo ritmo que los avances en la aproximación molecular para el
estudio de los receptores presinápticos y postsinápticos. El
patch-
clamping
es una técnica que emplea una micropipeta de cristal
para sondear la superficie de la membrana hasta que se
localiza un único receptor funcional. Se presiona la punta de la
pipeta en el interior de los lípidos de la membrana y se dispone el
aparato electrónico para mantener el potencial de membrana pinzado
(es decir, fijo) y medir la corriente que fluye a través del canal del
receptor. La solución en la pipeta puede contener acetilcolina, un
relajante muscular, otra sustancia, o una mezcla de fármacos. Pueden
monitorizarse los cambios eléctricos mediante la aplicación de estas
sustancias al receptor a través de la micropipeta.
4
© ELSEVIER. Fotocopiar sin autorización es un delito
Sección I
Fisiología y anestesia
Fisiología y farmacología neuromusculares
115
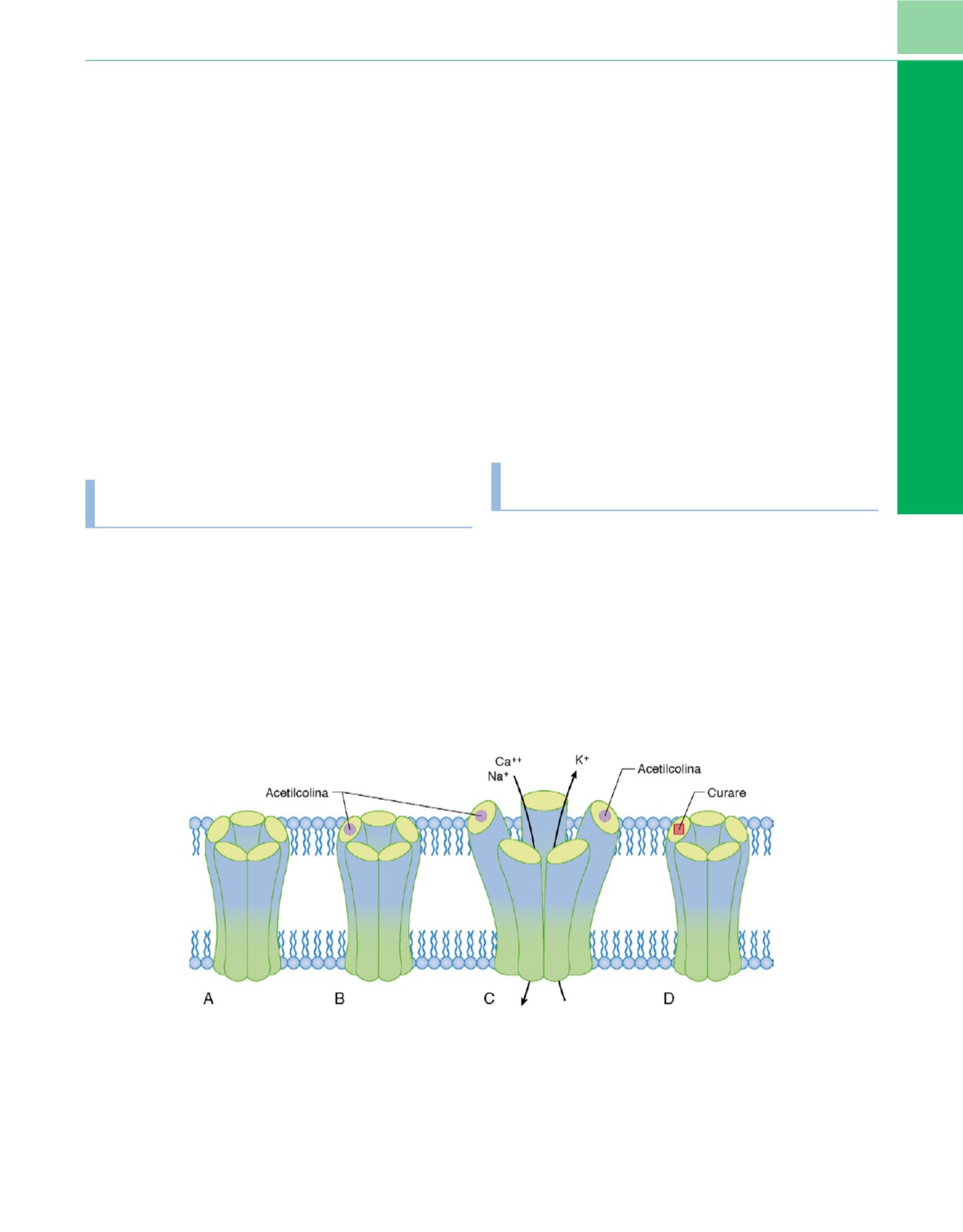
Figura 4-5
Acciones de la acetilcolina o el curare sobre los receptores de la placa terminal.
A,
El canal iónico está inactivo y no se abre en ausencia de
acetilcolina.
B,
Incluso la unión de una molécula de acetilcolina
(círculo relleno)
a uno de los dos sitios de unión no abre el canal.
C,
Cuando la acetilcolina se
une a los sitios de reconocimiento de ambas subunidades
a
simultáneamente
(círculos rellenos)
desencadena un cambio de conformación que abre el canal y
permite que los iones fluyan a través de la membrana.
D,
Acción de antagonistas como el curare
(cuadrado relleno)
. La acetilcolina compite con la
tubocurarina por el reconocimiento del lugar del receptor pero también puede reaccionar con la acetilcolinesterasa. La tubocurarina es un prototipo de
relajante muscular no despolarizante. La inhibición de la enzima aumenta la vida de la acetilcolina y la probabilidad de que reaccione con un receptor. Cuando
uno de los dos sitios de unión (reconocimiento) está ocupado por curare, el receptor no se abrirá, incluso si el otro lugar está ocupado por acetilcolina.