








 117 / 2894
117 / 2894

considerar agonistas a pesar del hecho de que bloqueen la
neurotransmisión después de una estimulación inicial. Desde el
punto de vista estructural, la succinilcolina consiste en dos molé-
culas de acetilcolina unidas. Por tanto, no es sorprendente que
pueda mimetizar los efectos de la acetilcolina. La succinilcolina o
el decametonio pueden unirse al receptor, abrir el canal, hacer
pasar corriente y despolarizar la placa terminal. Estos agonistas,
similares a la acetilcolina, se adhieren sólo brevemente; cada aper-
tura de un canal tiene una duración muy corta, 1 milisegundo o
menos. Sin embargo, la respuesta a la acetilcolina es del orden de
milisegundos debido a su rápida degradación por la acetilcolines-
terasa, y la placa terminal vuelve a su estado de reposo durante un
tiempo prolongado antes de que llegue otro impulso nervioso. En
cambio, los relajantes despolarizantes tienen de manera caracte-
rística una acción bifásica sobre el músculo, una contracción
inicial, seguida de una relajación que puede durar minutos u
horas. Los relajantes despolarizantes, puesto que no son suscepti-
bles a la hidrólisis por acetilcolinesterasa, no se eliminan de la
hendidura sináptica hasta que son aclarados del plasma. El tiempo
necesario para aclarar el fármaco del organismo es el principal
determinante de la duración del efecto del fármaco. El aclara-
miento corporal total del relajante es muy lento en comparación
con la acetilcolina, incluso cuando la colinesterasa plasmática es
normal. Puesto que las moléculas del relajante no se aclaran de la
hendidura con celeridad, reaccionan de forma rápida con los
receptores, adhiriéndose a uno casi inmediatamente después de
haberse separado de otro, y por tanto despolarizando repetidas
veces la placa terminal y abriendo los canales.
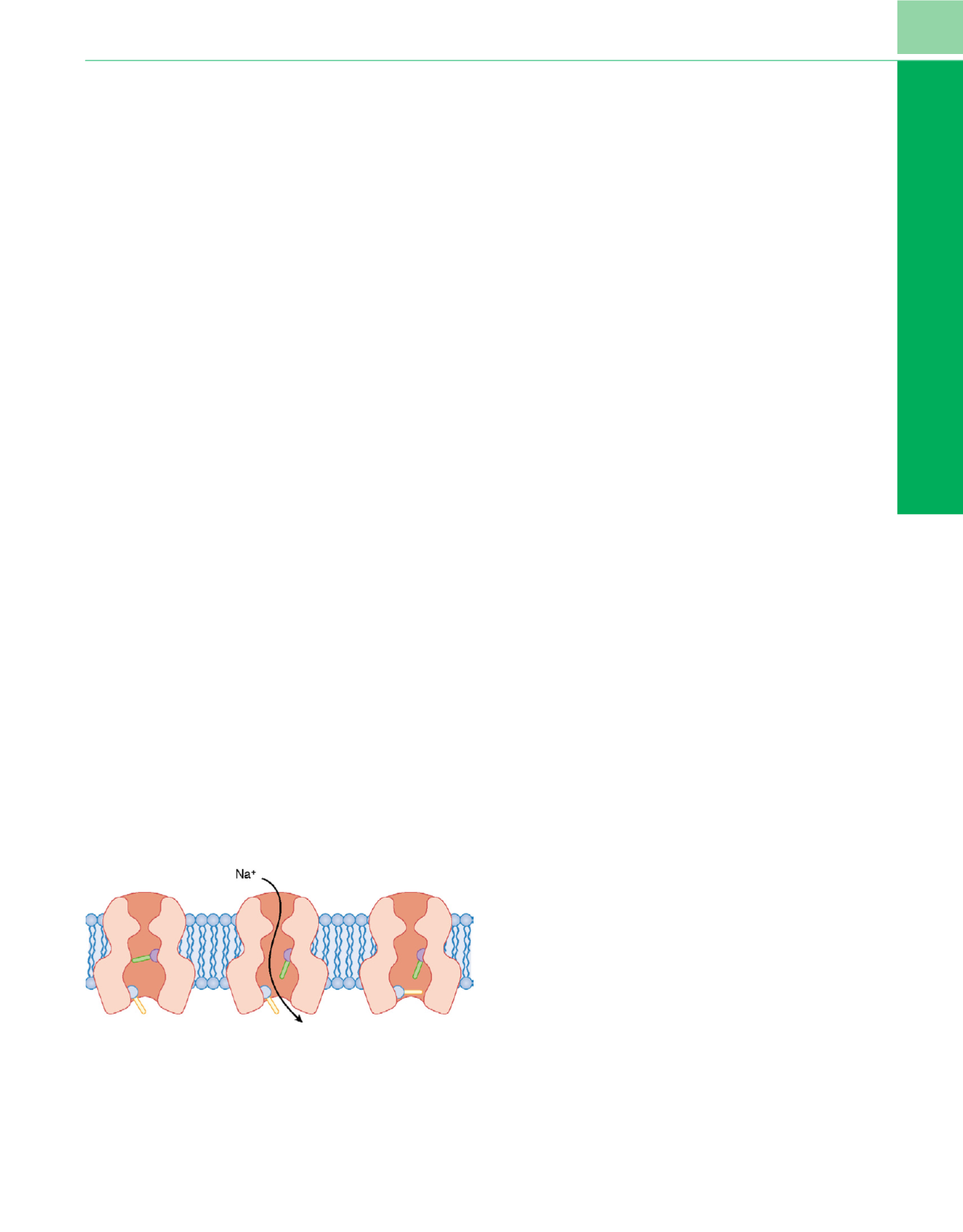
El desplazamiento rápido desde la excitación de la contracción
muscular hasta el bloqueo de la transmisión por los relajantes despo-
larizantes se produce porque la placa terminal está continuamente
despolarizada.Esto ocurre por la yuxtaposición en el borde de la placa
terminal sobre la membrana muscular de una clase distinta de canal
iónico: el canal de sodio, que no responde a agentes químicos, aunque
se abre cuando se expone a un cambio del voltaje transmembrana. El
canal de sodio también es una proteína cilíndrica transmembranosa
a través de la cual pueden fluir los iones de sodio. Dos partes de su
estructura actúan como puertas que permiten o detienen el flujo de
iones de sodio
69
.Ambas puertas deben estar abiertas para que el sodio
fluya a través del canal; el cierre de una de ellas corta el flujo. Puesto
que estas dos puertas actúan secuencialmente, un canal de sodio tiene
tres estados conformacionales funcionales y puede moverse de
manera progresiva de un estado a otro (fig. 4-6).
Cuando el canal de sodio se encuentra en situación de reposo,
la puerta inferior (es decir, la puerta de inactivación o dependiente
de tiempo) está abierta, pero la puerta superior (es decir, la puerta
dependiente de voltaje) está cerrada, y los iones de sodio no pueden
pasar. Cuando la molécula está sujeta a un cambio brusco de voltaje
por despolarización de la membrana adyacente se abre la puerta
superior y, puesto que la puerta inferior (dependiente de tiempo)
sigue abierta, el sodio fluye a través del canal. La puerta dependiente
de voltaje permanece abierta tanto tiempo como la molécula esté
sujeta a la influencia despolarizadora de la membrana de alrededor;
no se cerrará hasta que desaparezca la despolarización. Sin embargo,
poco tiempo después de que la puerta dependiente de voltaje se
abre, la puerta inferior se cierra y corta de nuevo el flujo de iones.
No puede abrirse de nuevo hasta que la puerta dependiente de
voltaje se cierre.Cuando cesa la despolarización de la placa terminal,
se cierra la puerta dependiente de voltaje; la puerta dependiente de
tiempo se abre y el canal de sodio vuelve a su situación de reposo.
El proceso completo es breve cuando la despolarización se produce
con acetilcolina. La respuesta inicial de un relajante muscular
despolarizante se parece a la de la acetilcolina, pero, dado que el
relajante no se hidroliza fácilmente, la despolarización de la placa
terminal no es breve.
La despolarización de la placa terminal por el relajante
determina al principio la apertura de la puerta mediada por voltaje
en los canales de sodio adyacentes, lo que produce una onda de
despolarización que barre a lo largo del músculo, provocando la
contracción muscular. Poco tiempo después de abrirse la puerta
dependiente de voltaje, se cierra la puerta dependiente de tiempo.
Puesto que el relajante no es retirado de la hendidura, la placa
terminal sigue despolarizándose. Como los canales de sodio inme-
diatamente adyacentes a la placa terminal están influidos por la
despolarización de la misma, sus puertas dependientes de voltaje
permanecen abiertas y sus puertas inactivadoras, cerradas. Dado
que el sodio no puede fluir a través de un canal que tiene cerrada
una puerta inactivadora, la membrana muscular perisináptica no
se despolariza. Cuando se detiene el flujo de iones a través de los
canales de sodio de la zona perisináptica porque se han cerrado las
puertas inactivadoras, los canales que se encuentran más abajo
(más allá de la región perisináptica) se liberan de la influencia
despolarizadora. En efecto, la zona perisináptica se convierte en un
tampón que protege al resto del músculo de los eventos que se
producen en la placa terminal. Por consiguiente, la membrana
muscular está separada en tres zonas: la placa terminal, que se
despolariza por la succinilcolina; la zona perisináptica, en la cual
los canales de sodio están congelados en un estado inactivo; y el
resto de la membrana muscular, en la que los canales de sodio están
en una situación de reposo. Dado que una descarga de acetilcolina
del nervio no puede sobrepasar los canales de sodio inactivados en
la zona perisináptica, la transmisión neuromuscular está bloqueada.
Este fenómeno también se denomina
acomodación.
Durante la
acomodación, cuando la sinapsis es inexcitable a través del nervio
(transmisor), la estimulación eléctrica directa del músculo causa
una contracción muscular porque los canales de sodio más allá de
la zona sináptica están en su situación de reposo excitable.
Los músculos extraoculares contienen fibras tónicas con
inervación múltiple y químicamente excitables a lo largo de la
mayoría de su superficie
20,21
.A pesar de su inervación, los músculos
oculares expresan tanto receptores maduros como fetales
20
. No se
produce la acomodación, por lo que estos músculos pueden sufrir
una contractura sostenida en presencia de succinilcolina.La tensión
así desarrollada fuerza al ojo contra la órbita y provoca, en parte,
el aumento de la presión intraocular producida por relajantes des-
polarizantes. También existe evidencia de que los músculos extrao-
culares contienenun tipo especial de receptor que no se desensibiliza
con la presencia continuada de acetilcolina u otros agonistas (como
se comenta más adelante)
21
. Se desconoce si es la subunidad
g
inmadura del RACh o la subunidad
a
7 del RACh la que desem-
peña una función en esta resistencia a la desensibilización en los
músculos oculares.
4
© ELSEVIER. Fotocopiar sin autorización es un delito
Sección I
Fisiología y anestesia
Fisiología y farmacología neuromusculares
117
Figura 4-6
Esquema del canal de sodio. Las
barras
representan partes de la
molécula que actúan como puertas. La
barra superior
es voltaje dependiente;
la
barra inferior
es tiempo dependiente. El lado izquierdo del dibujo
representa la situación de reposo. Una vez activada por un cambio de voltaje,
la molécula y sus puertas progresan como se ilustra (de izquierda a derecha)
.
Véase el texto para los detalles.