








 121 / 2894
121 / 2894

incluso durante la presencia continua de colina
81
, lo que aumenta la
posibilidad de que el potasio fluya desde el interior de la célula
(145mEq/l, aproximadamente) hacia el espacio extracelular, incluido
el plasma (4,5mEq/l), por su gradiente de concentración. La
a
-cono-
toxina GI química inhibe específicamente a los RACh convenciona-
les del músculo (maduros e inmaduros) pero no inhibe a los RACh
a
7. Los RACh
a
7 musculares son diferentes de los RACh
a
7 neuro-
nales (ganglios autonómicos y cerebrales) debido a que los primeros
no son inhibidos intensamente por la metil-licacotinina, un antago-
nista selectivo de los RACh
a
7 neuronales. Los RACh
a
7 expresados
en el tejido neuronal también se desensibilizan fácilmente con
colina
83
, una característica que contrasta con los RACh
a
7 muscula-
res, los cuales no se desensibilizan con colina
81
. El RACh en el
músculo tiene también una afinidad menor por sus antagonistas,
incluidos el pancuronio y la
a
-bungarotoxina; por consiguiente, las
concentraciones de estos fármacos que se requieren para bloquear
la despolarización inducida por agonista en los RACh
a
7 deben ser
más altas que en los RACh musculares convencionales (
a
1,
b
1,
d
,
ε
/
g
)
81
. En los RACh convencionales, incluso la unión de una de las
subunidades
a
1 por un antagonista produce una inactivación del
receptor debido a que la acetilcolina necesita ambas unidades
a
1 del
RACh para su activación. Sin embargo, en el RACh
a
7, incluso
aunque las tres subunidades estén unidas a un antagonista (p. ej., un
relajante muscular), las otras dos subunidades siguen estando dispo-
nibles para unirse al agonista y provocar la despolarización. Esta
característica puede ser la responsable de la resistencia de los RACh
a
7, en contraposición a los RACh convencionales, a los efectos blo-
queantes de fármacos como el pancuronio
2
.
Todavía no se ha estudiado la farmacología clínica del RACh
a
7 muscular, aunque su farmacología básica ayuda a comprender
la hiperpotasemia relacionada con la succinilcolina. La denervación
química o física del músculo no sólo provoca una regulación al alza
y cambios cualitativos (subunidad
ε→
subunidad
g
) en los RACh
sino también una regulación al alza de los RACh
a
7 en el músculo.
La succinilcolina, un análogo sintético de la acetilcolina que consiste
en dos moléculas de acetilcolina juntas, es capaz de despolarizar no
solo los RACh convencionales sino también los RACh
a
7 en el
músculo
83
. Además, el metabolito de la succinilcolina, la colina,
puede despolarizar los RACh
a
7 con poca desensibilización. Los
efectos despolarizantes de la succinilcolina y de la colina sobre los
RACh
a
7 regulados al alza pueden producir una fuga continua de
potasio intracelular y el anegamiento del líquido extracelular, incluido
el plasma, lo cual conduce a una hiperpotasemia.
Mantenimiento de las uniones
neuromusculares maduras
A diferencia de otras células, las células musculares son poco corrien-
tes en el sentido de que tienen gran cantidad –generalmente cientos–
de núcleos por célula. Cada uno de estos núcleos posee los genes para
fabricar ambos tipos de receptores. Múltiples factores, como la acti-
vidad eléctrica, la señalización por factor de crecimiento (p. ej., insu-
lina, agrina y neurregulinas) y la presencia o ausencia de inervación,
controlan la expresión de los tres tipos de isoformas del receptor
19,31
.
Esto se aprecia con mayor claridad en el embrión en desarrollo con-
forme se forma la unión neuromuscular.Antes de que sean inervadas,
las células musculares de un feto sólo sintetizan los receptores inma-
duros y los RACh
a
7, de ahí el término isoforma fetal para el primer
receptor
.
La síntesis está dirigida por prácticamente todos los núcleos
en la célula, y los receptores se expresan por toda la membrana de la
célula muscular (v. fig. 4-7). Conforme el feto se desarrolla y el
músculo es inervado, las células musculares comienzan a sintetizar la
isoforma madura de los receptores, los cuales son insertados
exclusivamente dentro del área de la (futura) placa en desarrollo. El
nervio libera varios factores de crecimiento que influyen sobre el
aparato sintetizador de los núcleos próximos. En primer lugar, los
factores inducidos por el nervio hacen que los núcleos subsinápticos
incrementen la síntesis de RACh. Luego, la actividad eléctrica indu-
cida por el nervio provoca una represión de los receptores en el área
extrasináptica. Los factores de crecimiento derivados del nervio, que
incluyen la agrina y la AIRA/neurregulina, determinan que los recep-
tores se agrupen en el área subsináptica y aceleren la expresión de la
isoforma madura (fig. 4-8)
62-65
. En condiciones asociadas a una resis-
tencia insulínica, parece que se produce una proliferación de RACh
más allá del área sináptica. Las condiciones en las que se ha observado
una resistencia insulínica (es decir, una disminución de la señaliza-
ción del factor de crecimiento) son la inmovilización, quemaduras y
denervación
84-86
. En estas condiciones se observa una regulación al
alza asociada a la expresión de isoformas inmaduras y de RACh
a
7
1-3
.
Por tanto, la señalización de agrina y de neurregulina puede ser
importante para la supresión de los RACh
a
7.
Antes de la inervación, los RACh están presentes por toda la
membrana muscular. Después de la inervación, se concentran cada
vez más en la membrana postsináptica, y en el nacimiento su pre-
sencia es prácticamente nula en el área extrasináptica. El proceso de
inervación progresa de forma un tanto lenta durante la vida fetal, y
madura durante la lactancia y la infancia
15,27,33
. Con el tiempo, dis-
minuye la concentración de los receptores inmaduros y éstos desa-
parecen de la parte periférica del músculo. En el músculo adulto
4
© ELSEVIER. Fotocopiar sin autorización es un delito
Sección I
Fisiología y anestesia
Fisiología y farmacología neuromusculares
121
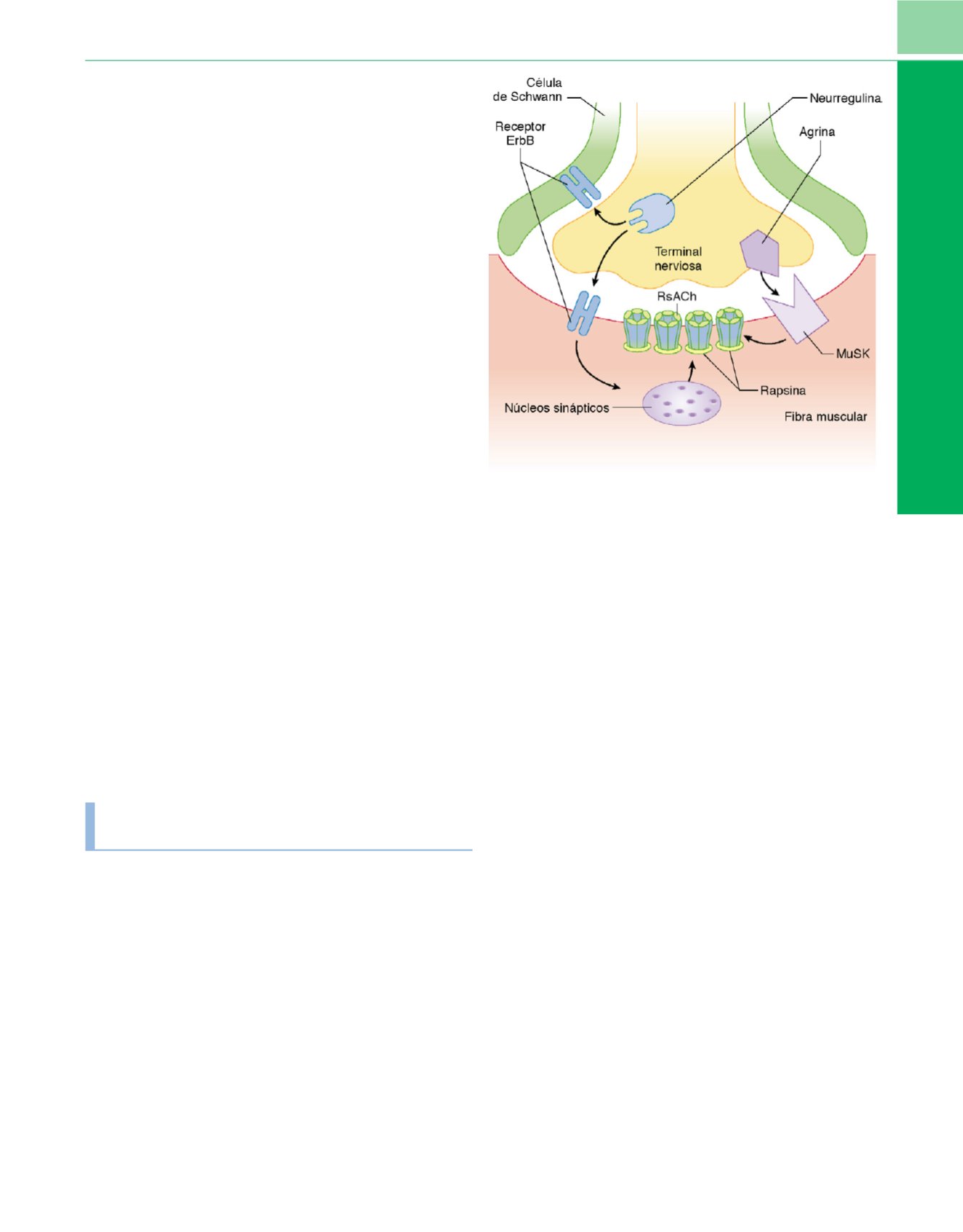
Figura 4-8
Diagrama de los acontecimientos dependientes de la agrina y
AIRA (actividad inducida por el receptor de acetilcolina) durante la
maduración de la unión neuromuscular. Después del establecimiento de un
nervio sobre el músculo, se liberan los factores de crecimiento, incluidas la
agrina y las neurregulinas. La señalización por neurregulina es esencial para la
supervivencia de la célula de Schwann, y las células de Schwann son
esenciales para el mantenimiento axonal. La interacción de la agrina con su
receptor MuSK (cinasa específica del músculo) favorece el agrupamiento de
proteínas sinápticas, incluidos los receptores de acetilcolina (RACh), rapsina y
receptores ErbB. La AIRA/neurregulina es el mejor candidato para la
implicación en la conversión del receptor inmaduro que contiene la
subunidad
g
en el receptor maduro (inervado) que contiene la subunidad
ε
,
que es la sinapsis específica y, por tanto, no está insertada en el área
extrasináptica.