








 120 / 2894
120 / 2894

adulta inervada se denomina receptor adulto, maduro o sináptico. Se
expresa otra isoforma de RACh cuando hay un descenso de la acti-
vidad muscular, como se observa en el feto antes de la inervación,
después de una lesión de motoneurona superior o inferior, quemadu-
ras o sepsis, o después de otros acontecimientos que provocan un
incremento del catabolismo proteico muscular, incluidas sepsis o
inflamación generalizada
1-3
. Para diferenciarlos de los receptores
maduros o sinápticos, la otra isoforma se denomina receptor de ace-
tilcolina inmaduro, extrasináptico o fetal. Alguna evidencia sugiere
que la isoforma inmadura no se observa en el catabolismo muscular
proteico ni en la atrofia que aparece en la malnutrición
79
. Las diferen-
cias en la estructura proteica de las isoformas producen importantes
variaciones cualitativas entre las respuestas de pacientes individuales
a los relajantes, y parecen ser responsables de algunos de los resulta-
dos anómalos que se observan cuando se administran relajantes a
personas concretas. Estas diferencias cualitativas en las isoformas
también pueden causar variaciones en la función del músculo
52,53
.
Además de sus composiciones estructurales, las dos isofor-
mas tienen otras características diferenciales
1-3,31
.A nivel molecular,
ambos tipos de receptor están formados por cinco subunidades (v.
fig. 4-4). El receptor sináptico maduro es un pentámero de dos
subunidades
a
y una subunidad
b
,
d
y
ε
de cada una de ellas. El
receptor inmaduro está formado por dos subunidades
a
y una
subunidad
b
,
d
y
g
; es decir que en el receptor inmaduro está pre-
sente la subunidad
g
en vez de la subunidad
ε
. Las subunidades
g
y
ε
difieren muy poco en su homología de aminoácidos pero las
diferencias son lo suficientemente grandes como para afectar a la
fisiología y farmacología del receptor y de su canal iónico. Aunque
las denominaciones sináptico y extrasináptico implican que cada
uno está localizado en las áreas sináptica o extrasináptica, esto no
es estrictamente correcto. Los receptores sinápticos están siempre
confinados a la región de la placa terminal (perisináptica) de la
membrana muscular. El receptor inmaduro o extrasináptico puede
expresarse en cualquier lugar de la membrana muscular. A pesar
del nombre extrasináptico, no están excluidos de la placa terminal.
Durante el desarrollo y en determinadas situaciones patológicas,
los receptores sinápticos y extrasinápticos pueden coexistir en el
área perisináptica de la membrana muscular (fig. 4-7).
A diferencia de los RACh del músculo convencional, que
constan de las subunidades
a
1,
b
1,
d
y
ε
/
g
descritas anteriormente,
RACh formados por unidades
a
7 han sido hallados recientemente
en el músculo esquelético durante el desarrollo y en la denerva-
ción
80,81
. La evidencia preliminar (Martyn JAJ, resumen de la ASA
2007) sugiere la existencia de una expresión aumentada de proteína
(mediante Western blot) de RACh
a
7 en el músculo después de
una lesión por quemaduras y en la inmovilización. Estos RACh
a
7
son canales homoméricos (es decir, formados por las mismas subu-
nidades) dispuestos como pentámeros (v. fig. 4-4). Se piensa que
los bolsillos de unión al ligando (fármaco) están formados por
caras negativas y positivas de las interfases de unión de la subuni-
dad
a
7. Como se esperaba, el agonista endógeno de la acetilcolina
se une al RACh
a
7, y cada una de las cinco subunidades tiene el
potencial de unir moléculas de acetilcolina o de succinilcolina
82,83
.
Otros agonistas, como la nicotina y la colina, y antagonistas, inclui-
dos los relajantes musculares, pancuronio, toxina de cobra y
a
-bungarotoxina, también se unen al RACh
a
7
81-83
.
Los RACh
a
7 en el músculo presentan unas características
funcionales y farmacológicas inusuales cuando se los compara con
los RACh musculares convencionales (
a
1,
b
1,
d
,
ε
/
g
) o los RACh
a
7
neuronales en el cerebro. La colina, un precursor y metabolito de la
acetilcolina (y de la succinilcolina), es un agonista extremadamente
débil del RACh muscular convencional aunque es un agonista com-
pleto del RACh
a
7 muscular; es decir, concentraciones de colina que
no abran canales del RACh convencional abrirán canales del RACh
a
7
81
. Es más, no se produce una desensibilización del RACh
a
7
120
Fisiología y anestesia
I
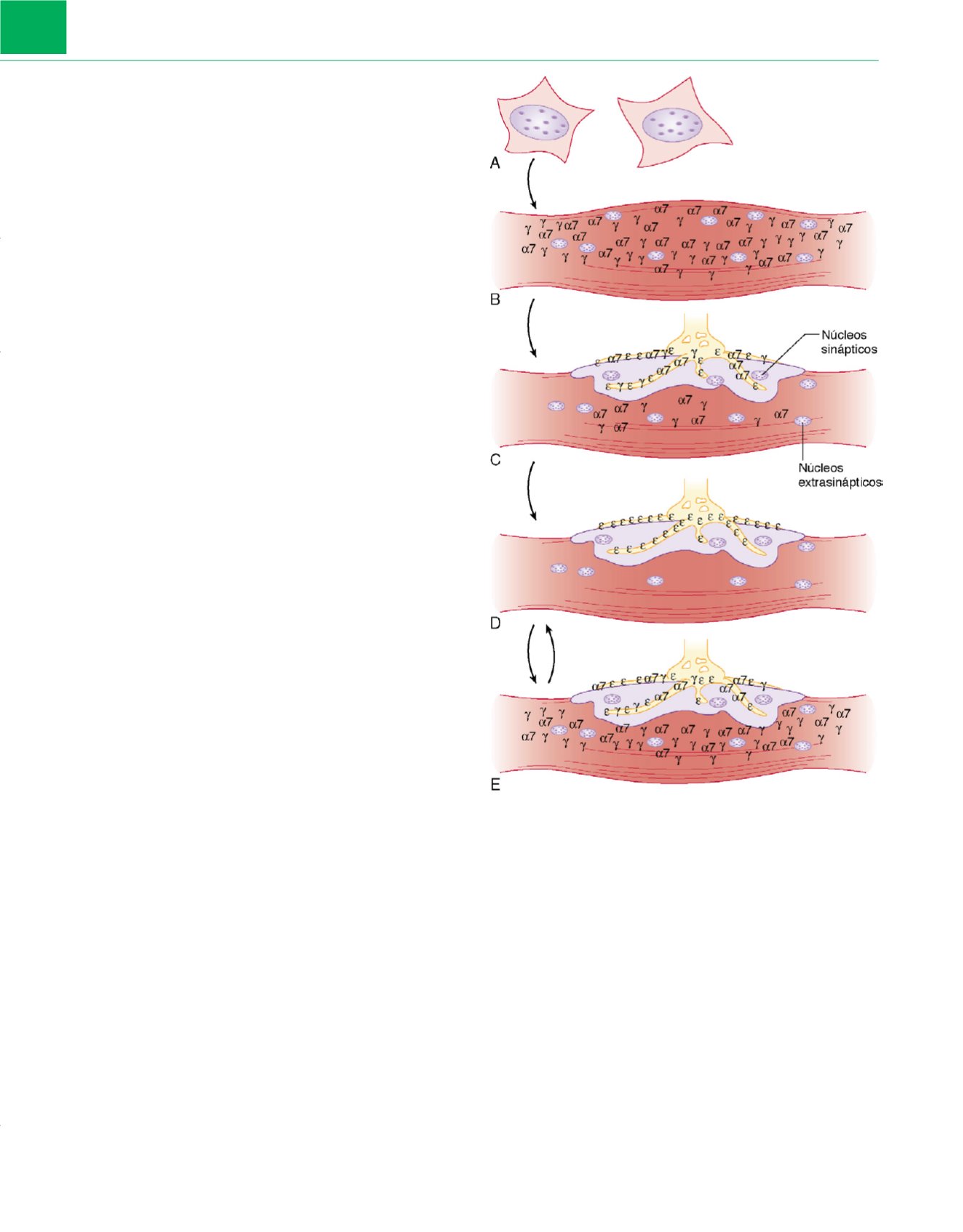
Figura 4-7
Distribución de los receptores de acetilcolina en el músculo
adulto en desarrollo, maduro, denervado, o en el músculo inmovilizado o
sujeto a catabolismo por inflamación.
A
y
B,
En la etapa fetal precoz, los
mioblastos mononucleados, derivados del mesodermo, se fusionan para
formar miotubos multinucleados. Los receptores de acetilcolina inmaduros
que contienen la subunidad
g
y los receptores de acetilcolina
a
7 neuronales
están diseminados por toda la membrana muscular antes de la inervación.
C,
Cuando el nervio contacta con el músculo, se produce el agrupamiento de
los receptores en la sinapsis y se asocia a la pérdida de algunos receptores
extrasinápticos.
D,
Se dice que se produce la maduración de la unión cuando
los receptores que contienen la subunidad
ε
reemplazan a los receptores que
contienen la subunidad
g
y
a
7. Incluso el miocito maduro es multinucleado,
aunque carece de receptores de acetilcolina extrasinápticos.
E,
La
denervación y algunos otros estados patológicos incluso sin denervación
anatómica (como quemaduras, inmovilización, tratamiento crónico con
relajantes musculares, ictus y sepsis) llevan a la reexpresión de los receptores
de acetilcolina con la subunidad
g
y
a
7 en las áreas sináptica y extrasináptica.
Los últimos cambios son potencialmente reversibles si la inmovilización/
catabolismo/inflamación muscular se restaura a la normalidad.