








 951 / 2894
951 / 2894

Otros cinco locus (17q21-24, 1q32, 3q13, 7q21-24 y 5p) se han
ligado a familias con pruebas de la contractura positivas y una
respuesta inusual a la anestesia, y se les ha designado locus PHM
2 a 6, respectivamente. Sin embargo, de estos cinco, el único gen
asociado a la HM es
CACNL1A3
207
, que codifica CaV1.1 (la subu-
nidad
a
1S
de DHPR), en el locus PHM3. Dos mutaciones causales
en este gen están ligadas a menos de un 1% de las familias con
PHM en todo el mundo. En algunos otros locus, todos los genes
dentro del locus se han excluido como causantes de la proclividad
a la HM. Debido a ello y por razones prácticas, el gen
RYR1
sigue
siendo la principal diana de los análisis genéticos clínicos
actuales.
Distribución de las mutaciones de
RYR1
Las mutaciones de cambio de aminoácido con PHM, ENC, o en
algunos casos ambos, están dispersas a través de la región codifi-
cadora del gen
RYR1
y todas permiten la transcripción de una
proteína que es supuestamente funcional
149,208
. Hasta hace poco se
pensaba que la mayoría de las mutaciones de
RYR1
estaba agru-
pada en tres «puntos calientes»: entre los aminoácidos 35 y 614
(región I de HM/ENC), entre los aminoácidos 2.163 y 2.458 (región
2 de HM/ENC) en la región pedicular mioplásmica de la proteína
y entre los aminoácidos 4.643 y 4.898 en el asa transmembranaria
terminal carboxílica o la región del poro (región 3 de PHM/ENC)
209
(fig. 27-4). Parece que la suposición de que había «puntos calientes»
se debía simplemente a un sesgo en el análisis de la muestra en la
medida que las mutaciones de cambio de aminoácido asociadas a
la HM o la ENC (o ambas) están dispersas sobre 54 de los 107 exones de
RYR1
. Aproximadamente el 41% de las mutaciones de HM
publicadas se encuentra en múltiples familias. Las mutaciones de
ENC se encuentran sobre todo en la región C terminal del gen
(exones 85 a 103) y sólo se han descrito 10 mutaciones (17%) en
más de una familia: R4861H (
n
=14),V4849I (
n
=9), I4898T (
n
=7),
L4824P (
n
=4), A4940T (
n
=4), G4638D (
n
=3), R4893W (
n
=3),
R4861C (
n
=2), R4893Q (
n
=2) y G4899E (
n
=2).
La HM y la ENC se han descrito en poblaciones occidenta-
les sobre todo, pero probablemente esto sea incorrecto y el resul-
tado de la manera y frecuencia con que se publican los casos. La
verdadera distribución étnica de la HM y la ENC es difícil de
asegurar. Parece que las mutaciones asociadas se agrupan en una
región dada del mundo, pero la distribución y frecuencia parecen
algo específicas de la población. En Reino Unido se han descu-
bierto 69 mutaciones de
RYR1,
25 de las cuales se encuentran en
una sola familia. G2434R se encuentra en alrededor del 40% de
las 434 familias con HM y mutaciones investigadas en Reino
Unido, y las siguientes mutaciones más frecuentes son T2206M
(10%) y G341R (8%). En Suiza, V2168M y I2336H son las muta-
ciones predominantes
210
, y en Alemania se han detectado R163C
(HM y ENC), R614C (HM), T2206M (HM), G2434R (HM) y
R2454H (HM) en cinco o más casos independientes
211,212
. G341R
y R614C son frecuentes en Francia
213
, y R614C se ha encontrado
también en varias familias con HM de Italia
213
y Canadá
214
. G341R
se ha encontrado con frecuencia en Bélgica
213
. La mutación común
a Europa y Norteamérica es G2434R, que ocurre en el 4 al 7% de
las familias europeas y el 5,5% de las norteamericanas
215
. Las
mutaciones de una sola familia son frecuentes en familias japone-
sas, chinas, taiwanesas, australianas y neozelandesas
216-218
, pero es
probable que esto pueda reflejar el pequeño número de casos
investigados allí. Como el cribado genético en estudios europeos
y norteamericanos se ha dirigido sobre todo a las regiones 1 y 2
de los «puntos calientes» originales del gen, la falta de mutaciones
de
RYR1
en algunas poblaciones estudiadas podría explicarse por
mutaciones de
RYR1
localizadas fuera de estas dos regiones o por
la afectación de otros genes.
Trastornos neuromusculares e hipertermia maligna
951
27
Sección
III
Control de la anestesia
© ELSEVIER. Fotocopiar sin autorización es un delito
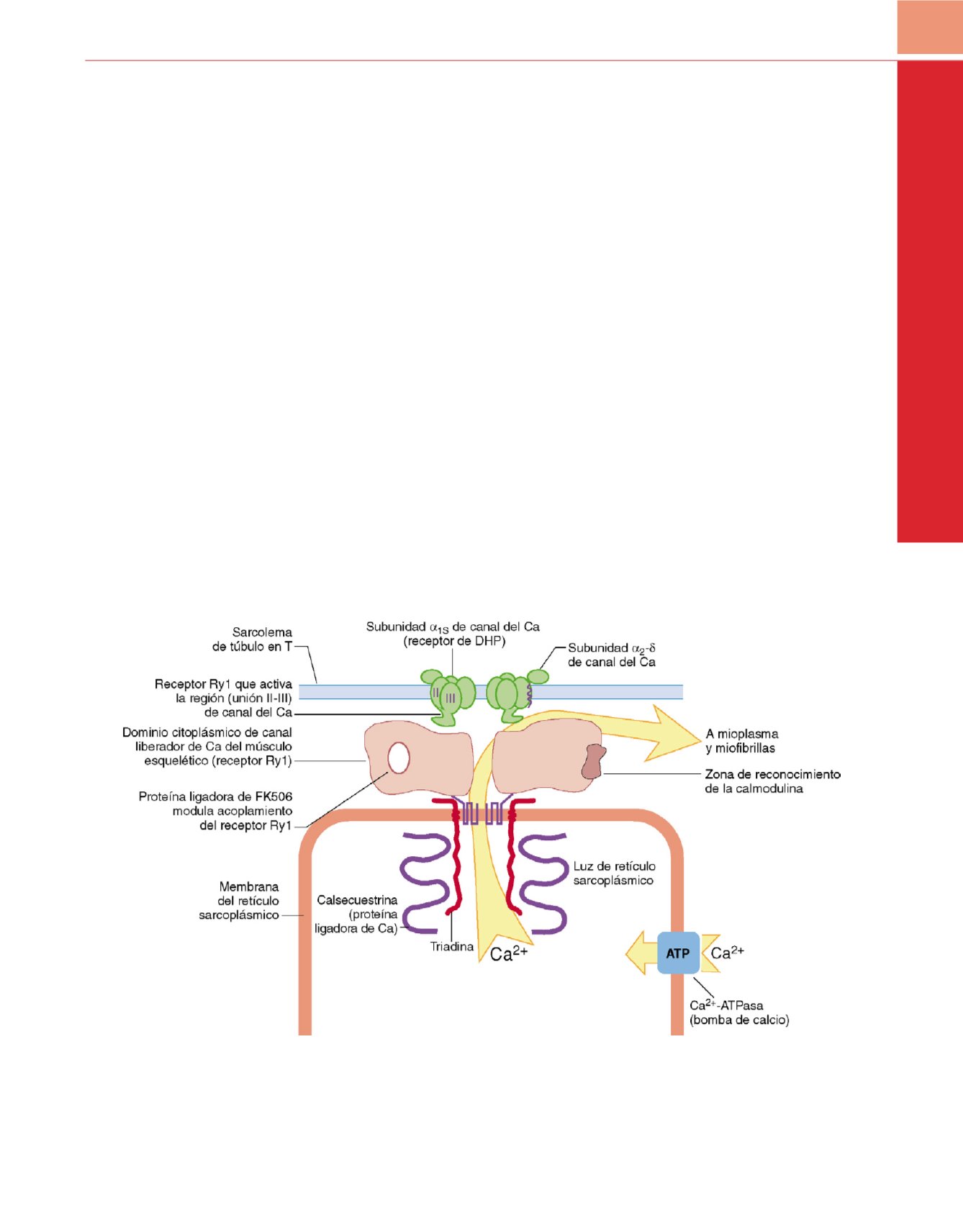
Figura 27-4
Representación esquemática de la unión tríada del músculo esquelético que muestra la proteína pedicular de la unión (receptor de rianodina
[Ry1]) y sus proteínas asociadas. En el músculo esquelético, la subunidad
a
1S
del receptor de la dihidropiridina (DHPR) participa en el acoplamiento de la
excitación y la contracción. Estos nexos físicos transmiten señales esenciales a través del espacio estrecho de la unión tríada que activan el receptor Ry1 y
liberan Ca
2+
del retículo sarcoplásmico.
(Adaptada de Pessah IN, Lynch C III, Gronert GA: Complex pharmacology of malignant hyperthermia.
Anesthesiology
84:1275, 1996.)