








 166 / 2894
166 / 2894

canales de liberación de calcio, sino que también forman las pro-
teínas de andamiaje que permiten el anclaje de muchas de las
principales proteínas reguladora
s 20.
Las mitocondrias se encuentran inmediatamente debajo del
sarcolema, introducidas entre las miofibrillas en el interior de la
célula. Contienen enzimas que favorecen la generación de trifos-
fato de adenosina (ATP), y son la central energética de los miocar-
diocitos. Además, las mitocondrias también pueden acumular
calcio y de esta forma contribuyen a la regulación de la concentra-
ción citosólica de calcio. Casi toda la información genética se
encuentra en el núcleo, de localización central. El citosol es el
microentorno lleno de líquido que hay en el interior del sarcolema,
excluyendo los orgánulos y el aparato contráctil y las proteínas.
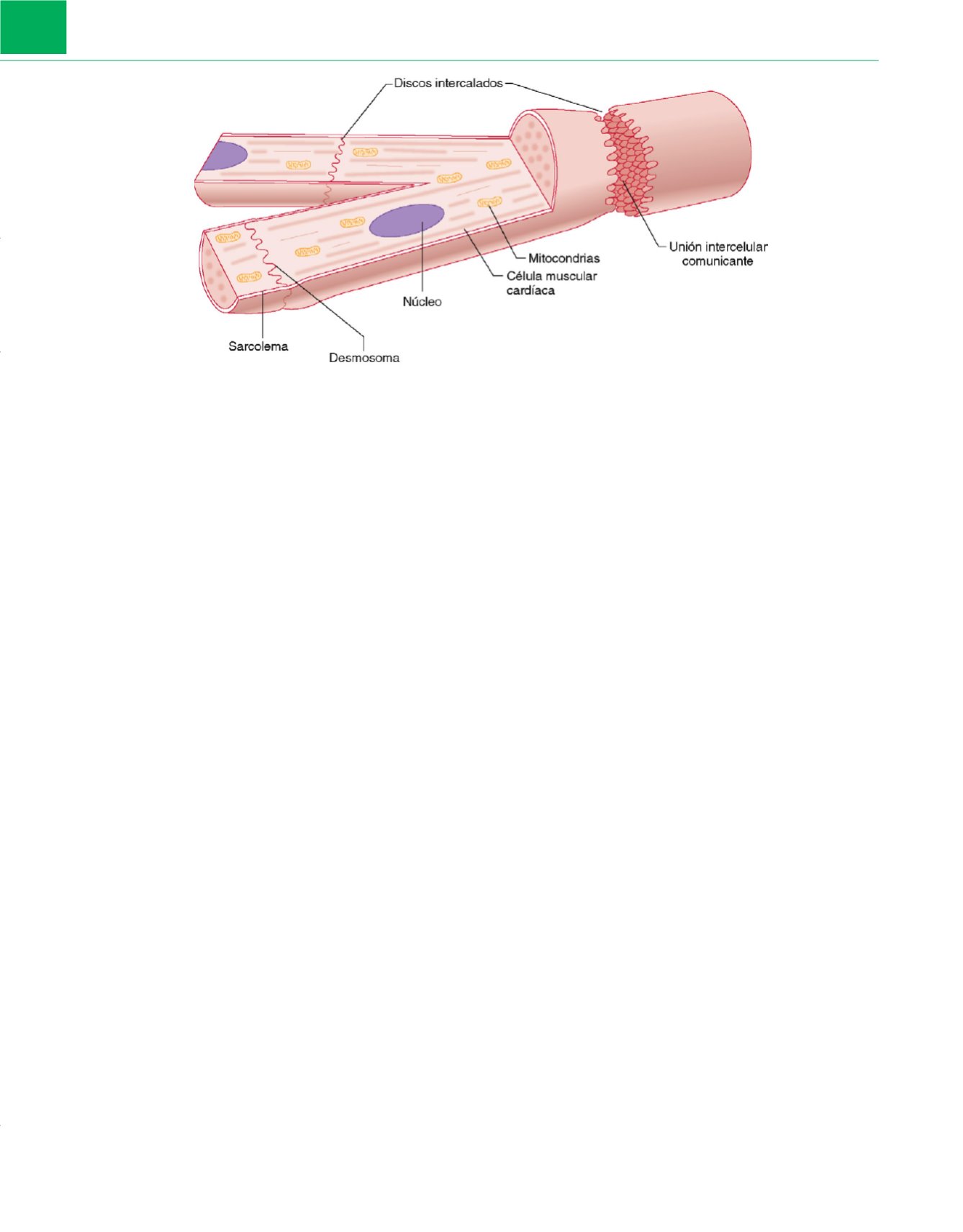
Las células musculares cardíacas contienen tres tipos de
uniones intercelulares: uniones intercelulares comunicantes
(gap
junctions),
desmosomas «puntuales», y desmosomas «en banda» (o
fascia adherente)
( fig. 6-10 ) 19,21 .Las uniones intracelulares comu-
nicantes son responsables del acoplamiento eléctrico y de la trans-
ferencia de moléculas pequeñas entre células, mientras que los
desmosomas ofrecen unión mecánica. Las zonas de adhesión for-
madas por los desmosomas «puntuales» anclan el citoesqueleto de
filamentos intermedios de la célula; las zonas formadas por la fascia
adherente anclan el aparato contráctil. Las uniones intercelulares
comunicantes están formadas por grupos de canales de la mem-
brana plasmática que unen directamente los compartimentos cito-
plásmicosdecélulasvecinas.Loscanalesdelasunionesintercelulares
comunicantes están formados por conexinas, una familia de pro-
teínas conservadas con múltiples genes. La principal isoforma de
las conexinas del corazón de mamíferos es la conexina 43; también
se expresan otras conexinas, sobre todo las conexinas 40, 45 y 37,
aunque en menores cantidade
s 20,21.
Los miocardiocitos de conducción, o células de Purkinje,
son células especializadas para la conducción de los potenciales de
acción propagados. Estas células tienen un contenido bajo de mio-
fibrillas y un núcleo prominente, y contienen muchas uniones
intracelulares comunicantes. Los miocardiocitos se pueden dividir
desde el punto de vista funcional en: 1) sistema de excitación,
2) sistema del AEC y 3) sistema contráctil.
Sistema de excitación
El potencial de acción celular que se origina en el tejido de con-
ducción especializado se propaga a las células individuales, en las
que inicia los fenómenos intracelulares que dan lugar a la contrac-
ción de la célula a través del sistema de excitación sarcolémico.
Potencial de acción
Los flujos iónicos a través de las membranas plasmáticas dan lugar
a despolarización (consecución de un potencial de membranamenos
negativo) y repolarización (consecución de un potencial de mem-
brana más negativo). Están mediados por proteínas de la membrana
con poros selectivos para determinados iones. Como estas proteínas
de los canales iónicos abren y cierran los poros en respuesta a los
cambios en el potencial de membrana, los canales están activados
por voltaje. Se ha observado que en el corazón canales de sodio,
potasio, calcio y cloruro contribuyen al potencial de acción.
Los tipos de potencial de acción del corazón se pueden
dividir en dos categorías: potenciales de acción de respuesta rápida,
que se encuentran en el sistema de His-Purkinje y en los miocar-
diocitos auriculares y ventriculares, y potenciales de acción de
respuesta lenta, que se encuentran en las células marcapasos de los
nódulos SA y AV. La
figura 6-11muestra un trazado típico de un
potencial de acción del sistema de His-Purkinj
e 12 .El gradiente
electroquímico del potasio a través de la membrana plasmática es
el determinante del potencial de membrana en reposo. Como con-
secuencia sobre todo de la entrada de Na
+
, el potencial de mem-
brana se despolariza, lo que da lugar a un ascenso muy rápido (fase
0). Cuando el potencial de membrana alcanza un nivel crítico, o
umbral, durante la despolarización, el potencial de acción se
propaga. Al ascenso rápido le sigue una repolarización transitoria
(fase 1). La fase 1 es un período de repolarización breve y limitada
que se debe principalmente a la activación de una corriente tran-
sitoria de salida de potasio, i
to
. La fase de meseta (fase 2) se produce
por la entrada neta de Ca
2+
a través de los canales de calcio de tipo L,
y por la salida de K
+
a través de diversos canales de potasio: la
corriente rectificadora de entrada i
k
, el rectificador tardío i
k1
, y
la i
to
. Se produce la repolarización (fase 3) cuando la salida de K
+
por
las tres corrientes de salida de potasio supera a la entrada de Ca
2+
,
lo que devuelve la membrana a su potencial de reposo. Se produce
muy poco flujo iónico durante la diástole (fase 4) en un potencial
de acción de respuesta rápida.
Por el contrario, durante la diástole (fase 4), las células mar-
capasos que tienen potenciales de acción de respuesta lenta tienen
capacidad de despolarización diastólica espontánea y generan el
ritmo cardíaco automático. Las corrientes de marcapasos durante
166
Fisiología y anestesia
I
Figura 6-10
El sarcolema que envuelve los miocardiocitos se especializa mucho para formar los discos intercalados, en los que los extremos de células
vecinas están en contacto. Los discos intercalados están formados por uniones intracelulares comunicantes y desmosomas «puntuales» y «en banda».