








 167 / 2894
167 / 2894

la fase 4 se deben a un aumento de las tres corrientes de entrada y
a una disminución de las dos corrientes de salida. Las tres corrien-
tes de entrada que contribuyen a la actividad espontánea del mar-
capasos incluyen dos mediadas por el calcio, la i
CaL
y la i
CaT
, y una
tercera que es una corriente catiónica mixta, la I
f
. Las dos corrientes
de salida son la corriente rectificadora tardía de potasio i
k
y la
corriente rectificadora de entrada de potasio i
k1
. En comparación
con el potencial de acción de respuesta rápida, la fase 0 es mucho
menos rápida, no hay fase 1, y la fase 2 es indistinguible de la fase 3
en el potencial de acción de respuesta lent
a 22.
Durante el potencial de acción cardíaco, el movimiento de
Ca
2+
hacia el interior de la célula y el movimiento de Na
+
fuera de
la célula crean un desequilibrio iónico. El intercambiador de Na
+
-
Ca
2+
restaura el equilibrio iónico celular mediante el transporte
activo de Ca
2+
fuera de la célula contra un gradiente de concentra-
ción, a la vez que introduce Na
+
en la célula por un mecanismo
dependiente de energía.
Acoplamiento excitación-contracción
El sarcolema, los túbulos transversos, el RS y los miofilamentos
( fig. 6-12 A ) 23son estructuras que participan en el AEC cardíaco. El
proceso del AEC comienza con la despolarización de la membrana
plasmática y la propagación de la excitación eléctrica por el sarco-
lema de los miocardiocitos. El calcio entra a través de canales de
la membrana plasmática que están concentrados en los túbulos T,
y dicha entrada activa la liberación de calcio desde el RS, lo que a
su vez estimula la contracción miofibrila
r 24 .El ubicuo segundo mensajero calcio es el principal actor del
AEC cardíaco (v.
fig. 6-12 B ) 23 .El reciclado del calcio en las estruc-
turas que participan en el AEC inicia y finaliza la contracción. La
activación del sistema contráctil depende de un aumento del calcio
citosólico libre y de su posterior unión a las proteínas contráctiles.
La entrada de calcio a través de los canales de calcio de tipo L
activados por voltaje (receptores dihidropiridínicos) produce un
pequeño aumento inicial del calcio intracelular. El aumento local
de la concentración de Ca
2+
como consecuencia de la entrada de
Ca
2+
a través de los canales de calcio de tipo L activados por voltaje
activa los canales de liberación de calcio (receptores de rianodina),
induce una liberación adicional de calcio desde las cisternas sub-
sarcolémicas del RS y, de esta forma, da lugar a un gran aumento
del calcio intracelular. Sin embargo, el aumento del calcio intrace-
lular es transitorio en la medida en que el calcio es retirado por:
1) la captación activa por la bomba de calcio-adenosina trifosfatasa
(ATPasa) del RS, 2) la salida de calcio desde el citosol por el inter-
cambiador de Na
+
-Ca
2+
y 3) la unión del calcio a proteína
s 25 .El RS constituye el armazón anatómico y es el principal
orgánulo para el reciclado del calcio. Es la localización de los depó-
sitos de calcio intracelulares. La liberación y recaptación cíclicas de
calcio por el RS regulan la concentración citosólica de calcio y
acoplan la excitación con la contracción. La proximidad física de
los canales de calcio de tipo L con los receptores de rianodina en
la membrana del RS facilita la liberación de calcio inducida por
calcio. La región del «pie» del receptor de la rianodina es la parte
que se extiende desde la membrana del RS hasta los túbulos T, en
los que están localizados los canales de calcio de tipo
L 16,25,26 .El RS también participa en la recaptación de calcio que inicia
la relajación o finaliza la contracción. La Ca
2+
-ATPasa sarcoplásmica/
del retículo endoplásmico (SERCA) es la bomba dependiente de
ATP que bombea activamente la mayor parte del calcio de nuevo
hacia el RS después de su liberación. La SERCA constituye cerca del
90% de todas las proteínas del RS, y en reposo es inhibida por la
fosfoproteína fosfolambano. El fosfolambano es una proteína de la
membrana del RS que es activa en la forma desfosforilada. La fosfo-
rilación por diversas cinasas como consecuencia de la estimulación
b
-adrenérgica o por otros estímulos inactiva el fosfolambano y libera
su acción inhibidora sobre la SERCA. Se produce retroalimentación
positiva, que da lugar a una fosforilación adicional del fosfolambano
y a una mayor actividad de la SERCA. Posteriormente, la recaptación
activa de calcio por la SERCA favorece la relajació
n 16,25,26 .Una vez captado por el RS, el calcio se almacena hasta que
se libera en el ciclo siguiente. La calsecuestrina y la calreticulina
son dos proteínas de almacenamiento del RS. La calsecuestrina es
una proteína de carga elevada que se localiza en el componente de
las cisternas del RS cerca de los túbulos T. Como está cerca de los
canales de liberación de calcio, el calcio almacenado se puede
descargar rápidamente para su liberación una vez que se han esti-
mulado los canales de liberación de calcio.
El calcio citosólico también se puede retirar por extrusión a
través de la bomba de calcio sarcolémica y por la actividad del
intercambiador de sodio-calcio. La proteína calmodulina es un
importante «sensor» y regulador del calcio intracelula
r 18.
Sistema contráctil
Elementos contráctiles
La unidad de trabajo básica de la contracción es el sarcómero, que
se define como la distancia entre dos líneas Z consecutivas (Z es la
abreviatura de la palabra alemana
Zuckung,
o «contracción»), que
unen en serie los sarcómeros. Cada sarcómero está formado por
una banda A central que está separada por la mitad de una banda I
de las líneas Z a ambos lados, porque la línea Z divide en dos
partes iguales la banda I. En la
figura 6-13se puede ver una repre-
sentación esquemátic
a 12. En cada sarcómero hay dos proteínas
contráctiles principales (v. la sección siguiente) y una proteína no
contráctil, la titin
a 25. Las dos proteínas contráctiles son la actina,
el filamento fino, y la miosina, el filamento grueso. Los filamentos
de actina y de titina están anclados a la línea Z, pero los filamentos
de miosina no llegan realmente a las líneas Z. La titina, la tercera
Fisiología cardíaca
167
6
Sección I
Fisiología y anestesia
© ELSEVIER. Fotocopiar sin autorización es un delito
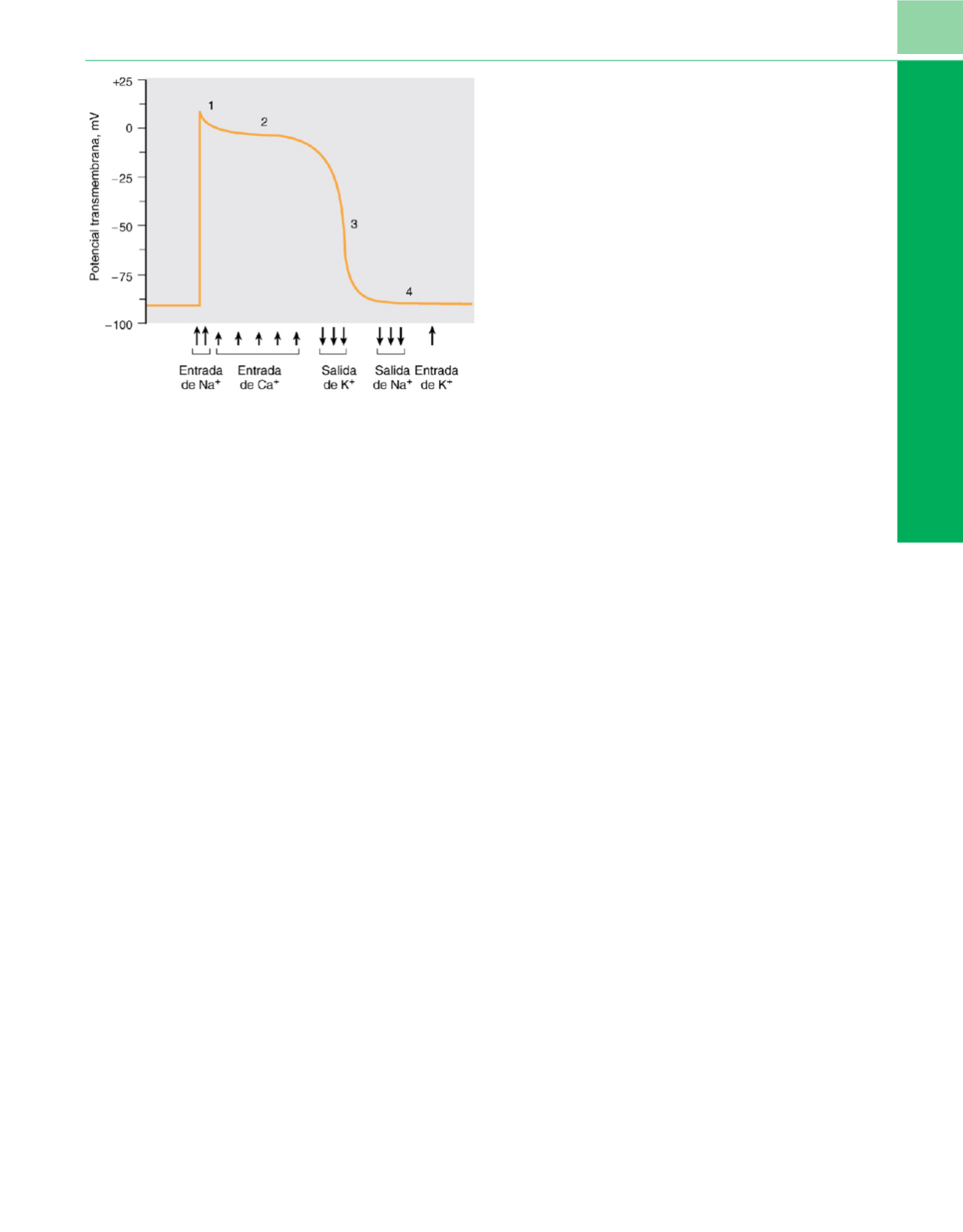
Figura 6-11
Fases de los potenciales de acción celulares, y principales
corrientes asociadas en los miocitos ventriculares. La espiga de la fase 0 inicial
y el sobreimpulso (1) están producidos por una corriente de entrada rápida de
Na
+
, la fase de meseta (2) por una corriente lenta de Ca
2+
a través de los
canales de Ca de tipo L, y la repolarización (fase 3) por corrientes de salida de
K
+
. La fase 4, el potencial de reposo (salida de Na
+
, entrada de K
+
), es mantenida
por la Na
+
-K
+
-ATPasa. El intercambiador de Na
+
-Ca
2+
es el principal responsable
de la salida de Ca
2+
. En el tejido del sistema de conducción especializado se
produce despolarización espontánea durante la fase 4, hasta que se alcanza el
voltaje que da lugar a la apertura del canal de Na.
(De LeWinter MM, Osol G:
Normal physiology of the cardiovascular system.
En
Fuster V [ed.]:
Hurst’s The
Heart,
10.ª ed., Nueva York, McGraw-Hill, 2001, págs. 63-94.)