








 165 / 2894
165 / 2894

periférica de O
2
. El consumo de O
2
es la cantidad de O
2
que llega
a los capilares pulmonares desde los alveolos (q
2
). Como
q
1
+q
2
=q
3
,
Q˙ (Capo
2
) + q
2
= Q˙ (Cvpo
2
)
q
2
= Q˙ (Cvpo
2
) − Q˙ (Capo
2
)
q
2
= Q˙ (Cvpo
2
) − Capo
2
Q˙ = q
2
/(Cvpo
2
− Capo
2
)
Así, si se conoce la concentración de O
2
en la arteria pulmo-
nar (Capo
2
), la concentración de O
2
en la vena pulmonar (Cvpo
2
)
y el consumo de O
2
(q
2
), se puede determinar el gasto cardíaco.
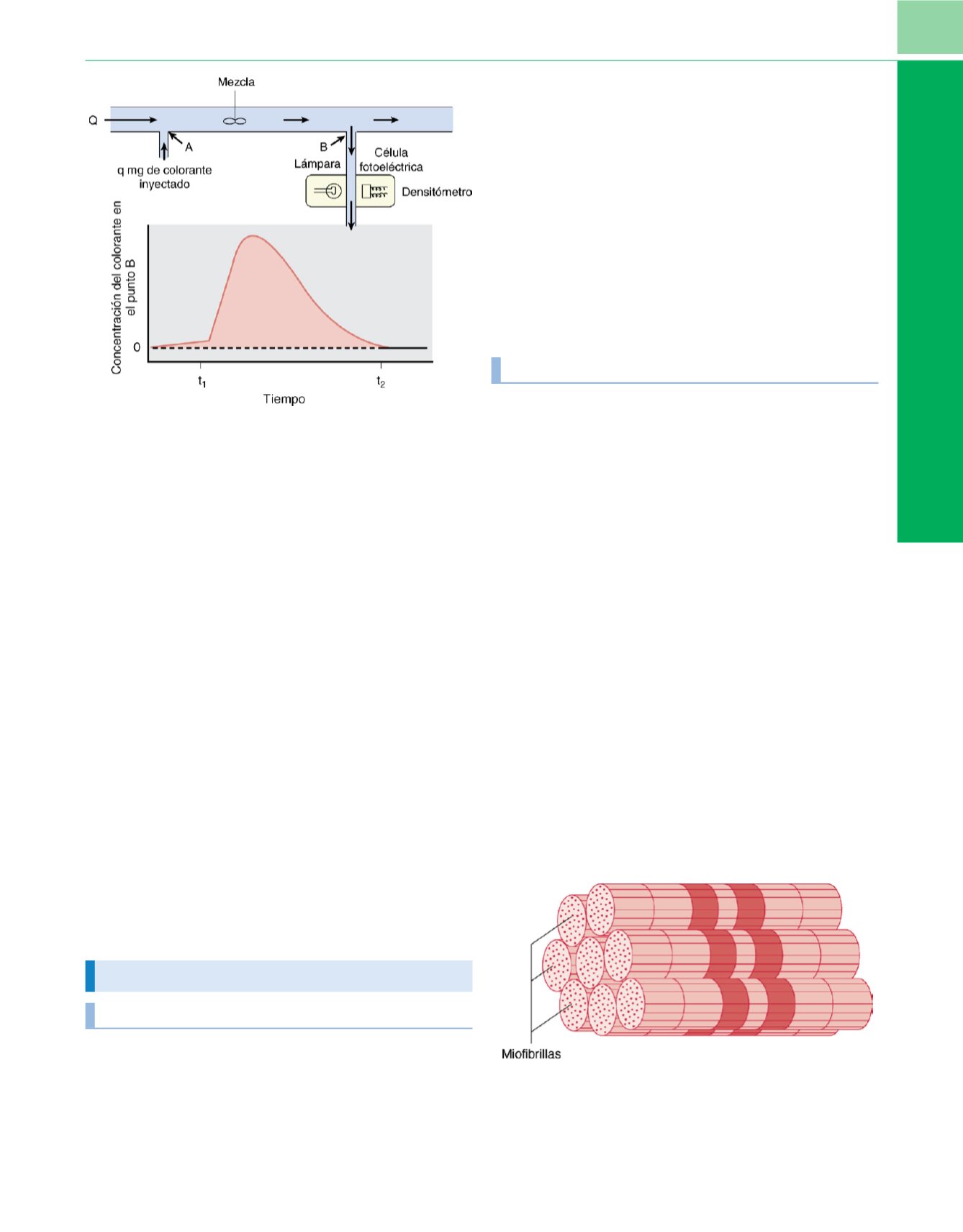
La técnica de dilución de marcadores es otro método que se
usa para determinar el gasto cardíaco, y también se basa en la ley
de conservación de la masa. Las dos técnicas de dilución de mar-
cadores que se utilizan con más frecuencia son los métodos de
dilución de colorantes y de termodilución. La
figura 6-8ilustra los
principios del método de dilución de colorante
s 1 .Fisiología cardíaca celular
Anatomía celular
A nivel celular, el corazón está formado por tres componentes
principales: tejido muscular cardíaco (miocardiocitos contrácti-
les), tejido de conducción (células de conducción) y tejido conjun-
tivo extracelular. Un grupo de miocardiocitos con su red de
soporte de tejido conjuntivo o matriz extracelular forma una mio-
fibra
( fig. 6-9). Las miofibras adyacentes están conectadas por
bandas de colágeno. La matriz extracelular es el producto sintético
de los fibroblastos y está formada por colágeno, que es el principal
determinante de la rigidez del miocardio, y otras importantes
proteínas de la matriz. Una de las proteínas de la matriz, la elas-
tina, es el principal componente de las fibras elásticas. Éstas son
en parte responsables de las propiedades elásticas del miocardi
o 6.
Otras proteínas de la matriz incluyen las glucoproteínas o proteo-
glucanos y las metaloproteínas de la matriz. Los proteoglucanos
son proteínas con cadenas de azúcares cortos, e incluyen sulfato
de heparano, condroitina, fibronectina y laminina. Las metalopro-
teínas de la matriz son enzimas que degradan el colágeno y otras
proteínas extracelulares. El equilibrio entre la acumulación de pro-
teínas de la matriz extracelular mediante síntesis y su degradación
por las metaloproteínas de la matriz contribuye a las propiedades
mecánicas y a la función del corazó
n 6.
Estructura y función de los miocardiocitos
Los miocardiocitos contráctiles individuales son células grandes de
entre 20
m
m (miocardiocitos auriculares) y 140
m
m (miocardiocitos
ventriculares) de longitud. Aproximadamente el 50% del volumen
celular de un miocardiocito contráctil está formado por miofibri-
llas, y el resto son mitocondrias, núcleo, retículo sarcoplásmico
(RS) y citosol. La miofibrilla es un haz similar a un bastón que
forma los elementos contráctiles del interior de los miocardiocitos.
En cada elemento contráctil hay proteínas contráctiles, proteínas
reguladoras y proteínas estructurales. Las proteínas contráctiles
suponen en torno al 80% de las proteínas miofibrilares, y el resto
son proteínas reguladoras y estructurale
s 16,17. La unidad básica de
contracción es el sarcómero, que se describirá más adelante.
El sarcolema, o membrana plasmática externa, separa el
espacio intracelular del extracelular. Rodea al miocardiocito y se
invagina hacia las miofibrillas a través de una red tubular extensa
conocida como túbulos transversos o túbulos T, y también forma
uniones intercelulares especializadas entre célula
s 18,19.
Los túbulos transversos o túbulos T están muy próximos a
un sistema intramembranario y al RS, que tiene una función
importante en el metabolismo del calcio que es crítico para el
acoplamiento excitación-contracción (AEC) del miocardiocito. El
RS se puede subdividir en RS longitudinal (reticular) y el RS de las
uniones. El RS longitudinal participa en la captación de calcio para
el inicio de la relajación. El RS de las uniones contiene grandes
canales para la liberación de calcio (receptores de rianodina) que
liberan los depósitos de calcio del RS como respuesta a la entrada de
calcio estimulada por la despolarización a través de los canales
de calcio sarcolémicos. Los receptores de rianodina no sólo son
Fisiología cardíaca
165
6
Sección I
Fisiología y anestesia
© ELSEVIER. Fotocopiar sin autorización es un delito
Figura 6-8
Ilustración que muestra el principio de la determinación del gasto
cardíaco con la técnica de dilución de marcadores. Este modelo asume que
no hay recirculación. Se inyecta una cantidad conocida de un colorante (q) en
el punto A de una corriente que fluye con un flujo Q˙ (ml/min). Se extrae una
muestra mixta del líquido que fluye más allá del punto B a una velocidad
constante a través de un densitómetro. En una curva se representa el cambio
de la concentración del colorante a lo largo del tiempo. El flujo se puede
medir dividiendo la cantidad de marcador inyectado en dirección proximal
por el área bajo la curva de concentración en dirección distal.
(De Berne RM,
Levy MN: The cardiac pump.
En Cardiovascular Physiology,
8.
a
ed. St. Louis,
CV Mosby, 2001, págs. 55-82.)
Figura 6-9
Organización de los miocardiocitos. El 50% del volumen del
miocardiocito está formado por miofibrillas; el resto son mitocondrias,
núcleo, retículo sarcoplásmico y citosol.