








 11 / 2894
11 / 2894

durante el sueño y activos durante la vigilia envían señales direccio-
nales que pueden ser inhibidoras mutuamente, excitadoras mutua-
mente o una inhibidora en una dirección con un retorno excitador,
es necesario estudiar de forma empírica los efectos reales de los anes-
tésicos sobre la salida neta del circuito porque los modelos existentes
no tienen en consideración toda la complejidad de los circuito
s 58 .Centros talámicos
Las teorías pasivas del sueño que formuló Bremer son similares a
muchos conceptos pasivos de la anestesia general. Un aspecto central
del sueño NREM y de la anestesia es que la corteza queda privada de
aferencia sensitiva.Ya sea por lesiones exógenas, como en el gato con
cerebro aislado de Bremer,o por el cierre endógeno de las compuertas
talámicas, los anestésicos parecen actuar sobre los circuitos del sueño
NREM,dando así lugar amecanismos de acción compartidos.Dentro
del tálamo hay una arquitectura sencilla de tipos celulares formada
por neuronas reticulares y neuronas talamocorticales que se comu-
nican con la corteza, a la vez que también integran las aferencias
periféricas
( fig. 1-8 ). La activación de las neuronas reticulares durante
el sueño NREM y la anestesia produce hiperpolarización de las neu-
ronas de interconexión talamocortical, lo que a su vez bloquea la
propagación del potencial de acción en todas las neuronas de inter-
conexión talamocorticales. En consecuencia, se impide que las neu-
ronas talamocorticales transmitan las aferencias periféricas hacia los
centros corticales superiores. Éste es el mecanismo mediante el cual
las compuertas talámicas se cierran para aislar, de forma transitoria
pero reversible, a la corteza de la periferi
a 25,59,60 .Se piensa que los
núcleos talámicos de la línea media tienen una participación funda-
mental en la generación de la conciencia consciente y la recepción
adecuada de las aferencias que proceden de la mayoría de los centros
activadores reticulares promotores del alertamient
o 21 .Los estudios de
imagen confirman una reducción selectiva a nivel regional del flujo
sanguíneo, el metabolismo y, por extensión, la actividad del tálamo
en la línea medi
a 49,61. Recientemente se ha fortalecido el respaldo a
un «interruptor» talamocortical de la consciencia ante el hallazgo de
que la microinyección de nicotina en el núcleo centromedial del
tálamo revierte la hipnosis inducida por sevoflurano (se comenta más
adelante). Estas conclusiones están mitigadas por el hecho de que la
administración de nicotina en el núcleo centromedial produce con-
vulsiones. Sin embargo, el respaldo a la idea del tálamo central como
centro de activación que es capaz de revertir la inconsciencia también
procede de textos sobre el estado vegetativo persistente. La estimula-
ción de frecuencia elevada del tálamo central en la rata se ha asociado
a activación cortical generalizada y mejora de la función cognitiv
a 62 .Además, se ha demostrado que la estimulación cerebral profunda del
tálamo central revierte algunos de los déficits conductuales en un
paciente que ha tenido una lesión cerebral traumátic
a 63 .Como los receptores nicotínicos de acetilcolina se expresan
mucho en el tálamo, y como muchos anestésicos inhiben la trans-
misión de señales a través de los receptores nicotínicos de acetilco-
lina, la supresión del sistema de activación colinérgico puede ser un
mecanismo mediante el cual muchos anestésicos producen incons-
cienci
a 64. Las medidas de EEG procesadas de la profundidad de los
anestésicos también muestran la importante participación del
sistema activador colinérgico, porque las infusiones intracerebro-
ventriculares de neostigmina o del agonista muscarínico oxotremo-
rina despiertan a ratas que han sido anestesiadas con isofluran
o 65 .Centros hipotalámicos
Los núcleos talámicos reciben aferencias procedentes del sistema
activador reticular ascendente del tronco encefálico y también
reciben aferencias hipotalámicas de los centros activos durante la
vigilia, como las neuronas histaminérgicas y orexinérgicas (v.
fig. 1-5 ). Como ya se ha señalado, las compuertas talámicas se cierran
durante el sueño NREM y la exposición a varios anestésicos, y este
cierre se ve facilitado por la disminución de la llegada de señales
monoaminérgicas, colinérgicas y orexinérgicas durante la anestesia.
Los anestésicos GABAérgicos, como el propofol y los barbituratos,
ejercen sus efectos hipnóticos mediante la inactivación de las neu-
ronas histaminérgicas del NT
M 66(v.
fig. 1-4 ). Esta acción se puede
explicar a nivel molecular por la potenciación de la proyección
GABAérgica inhibidora que procede del centro del sueño, el núcleo
POVL. A su vez, la desinhibición del núcleo POVL «apaga» otros
grupos activos durante la vigilia y refuerza aún más la actividad del
núcleo POVL. Este mecanismo de proalimentación estabiliza el
estado hipnótic
o 14 .El bloqueo de la señal histaminérgica promotora
de la vigilia también es el mecanismo mediante el cual el fármaco
antihistaminérgico difenhidramina precipita el sueño. La recupera-
ción o la salida de la hipnosis anestésica está facilitada por las
neuronas orexinérgicas promotoras de la vigilia, que son inhibidas
por anestésicos volátiles como el isoflurano y el sevofluran
o 67.
Sueño, memoria y consciencia
11
1
Sección I
Fisiología y anestesia
© ELSEVIER. Fotocopiar sin autorización es un delito
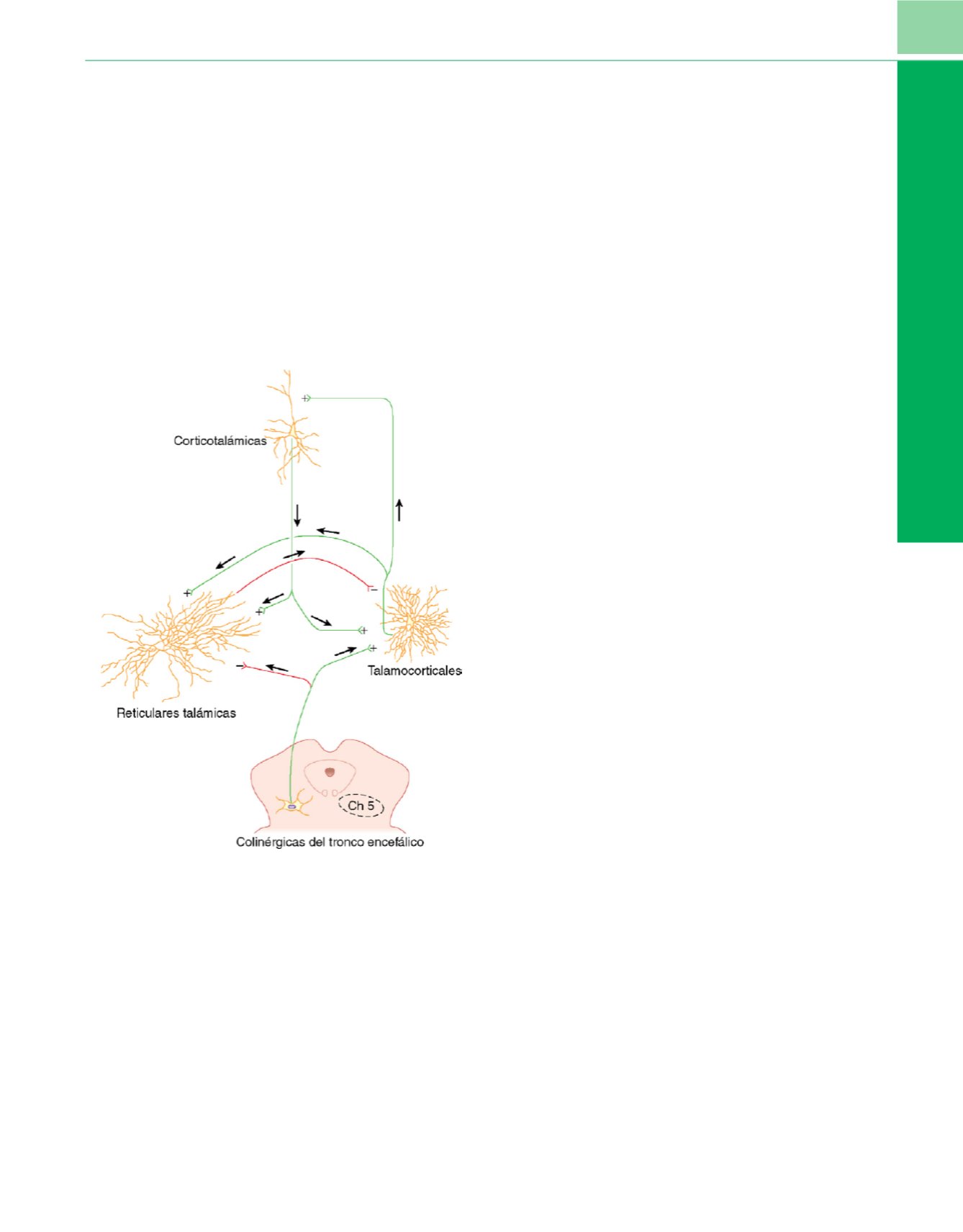
Figura 1-8
La transición desde la vigilia hasta el sueño no de movimientos
oculares rápidos (NREM) se asocia a cambios característicos en el
electroencefalograma que se correlacionan con cambios subyacentes de los
patrones de descarga eléctrica de los sistemas corticotalámicos. Durante el
sueño NREM las neuronas talamocorticales son hiperpolarizadas por las neuronas
reticulares talámicas (
axón verde
superior). Esta acción impide que las señales
periféricas entrantes se transmitan hacia las neuronas corticotalámicas de la
corteza y produce de forma transitoria pero eficaz desaferentación de la corteza.
Las aferencias colinérgicas hacia el tronco encefálico pueden hiperpolarizar
simultáneamente las neuronas reticulares talámicas a la vez que despolarizan las
neuronas talamocorticales, devolviendo así el potencial de membrana de
las neuronas talamocorticales al valor inicial y restaurando la propagación del
potencial de acción que transfiere las aferencias sensitivas periféricas hacia las
neuronas corticotalámicas. Estudios recientes indican que cambios similares de
las propiedades de descarga de los bucles corticotalámicos pueden subyacer
también a la hipnosis inducida por anestésicos. Las conexiones excitadoras se
muestran en
rojo,
con
signos de más
en las sinapsis. Las conexiones inhibidoras
se muestran en
verde,
con
signos de menos
en las sinapsis.
(Modificada de
Steriade M: The corticothalamic system in sleep.
Front Biosci
8:d878, 2003.)