








 227 / 2894
227 / 2894

neurohipófisis, donde se almacena en gránulos. La estimulación
nerviosa de los cuerpos celulares desencadena la exocitosis de la
AVP desde sus vesículas terminales hacia la circulación.
La AVP actúa sobre receptores-V
2
específicos en los tubos
colectores para inducir reabsorción de agua y disminución de la
diuresis, con una orina concentrada. También aumenta la reabsor-
ción de NaCl en el asa gruesa ascendente de Henle hacia el inters-
ticio medular, lo que mantiene su hipertonicidad y facilita la salida
del agua del tubo colector a lo largo de un gradiente osmótico. Esta
reabsorción origina la conservación tubular de agua y la retención
de agua libre (es decir, aclaramiento negativo de agua libre). El
efecto neto es que la AVP aumenta la osmolalidad urinaria y dis-
minuye la plasmática sin una alteración significativa de la excre-
ción de solutos.
El receptor-V
2
de la membrana basolateral celular del tubo
colector responde a la AVP a través de un mecanismo receptor
análogo al receptor
b
-adrenérgic
o 51. Mediante la activación de
una adenililciclasa acoplada a proteína G, el ATP se convierte en
monofosfato de adenosina cíclico (AMPc), que a su vez activa una
proteincinasa que provoca la migración de las vesículas prefor-
madas que contienen canales de agua acuaporina-2 y su fusión
con la membrana apical celular. Esta acción aumenta de forma
considerable la permeabilidad de la membrana al agua, que se
reabsorbe hacia la célula y desde allí al capilar peritubular. La
semivida plasmática de la AVP es de 5-15 minutos y este proceso
se invierte con rapidez cuando la concentración plasmática de
AVP disminuye.
Regulación de la secreción de AVP
Los osmorreceptores hipotalámicos son sensibles a aumentos tan
pequeños de la osmolalidad sérica como de un 1% por encima de
lo normal. Como se ilustra en la
figura 8-21A, el umbral para la
secreción de AVP (y de la sensación de sed) se encuentra entre 280 y
290mOsm/kg. Una vez superado dicho umbral, la tasa de secreción
muestra un aumento muy pronunciad
o 52 .Incluso una deshidrata-
ción leve produce una rápida antidiuresis y la osmolalidad urinaria
puede aumentar de 300 a 1.200mOsm/kg a medida que los niveles
plasmáticos de AVP suben de 0 a 5pg/ml (v.
fig. 8-21B).
Los descensos del volumen intravascular también estimulan
la secreción de AVP mediada por receptores de estiramiento con
aferencias vagales en la aurícula izquierda y las venas pulmonares.
La secreción de AVP inducida por hipovolemia supera las respues-
tas osmolares y contribuye al síndrome perioperatorio de secreción
inapropiada de hormona antidiurética (SIADH): retención de líqui-
dos, hipoosmolalidad e hiponatremia. La situación se agrava por la
administración de grandes cantidades de soluciones hipotónicas
que disminuyen la osmolalidad sérica. El estrés psíquico, a través
de conexiones corticales, también induce la liberación de AVP y
puede sobrepasar a los sensores osmóticos y de volumen.
El desencadenante más potente, con gran diferencia, para la
liberación de AVP es la hipotensión arterial sistémica mediada por
barorreceptores aórticos y carotídeos. Supera a todos los demás
desencadenantes y la concentración plasmática deAVP puede alcan-
zar niveles de 10 a 1.000 veces mayores de lo normal (v.
fig. 8-21C).
A estas concentraciones, la AVP actúa como un vasoconstrictor,
Fisiología renal
227
8
Sección I
Fisiología y anestesia
© ELSEVIER. Fotocopiar sin autorización es un delito
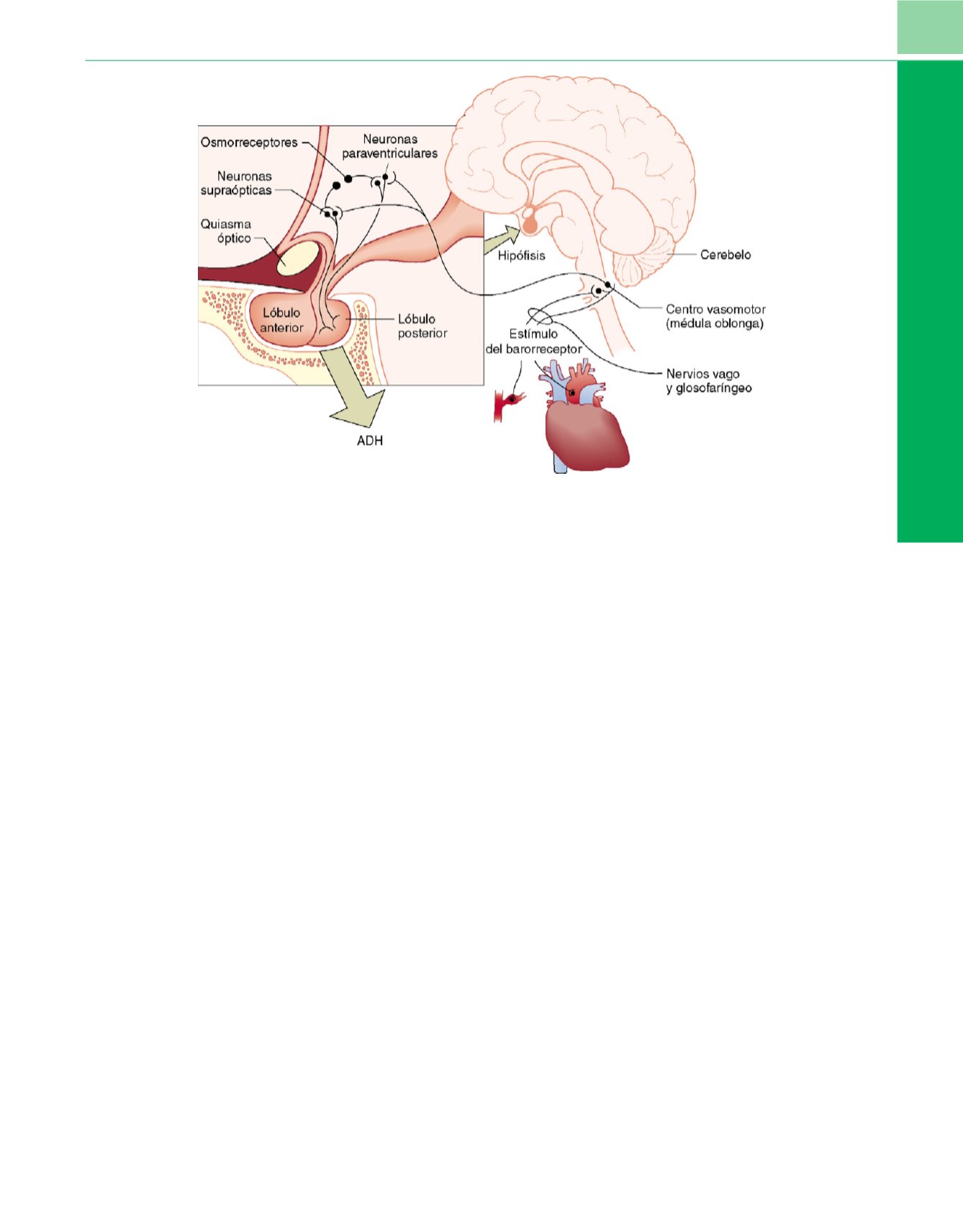
Figura 8-20
Síntesis y control de la arginina vasopresina (AVP).
Control osmótico.
Los osmorreceptores de las neuronas supraópticas y paraventriculares en el
hipotálamo detectan el aumento de la osmolalidad sérica. Los gránulos de AVP son trasportados por el axón tras su elaboración hacia el lóbulo posterior de la
hipófisis. Desde allí liberan a la circulación para alcanzar los receptores V
2
en el túbulo distal, donde éstos retienen agua y normalizan la osmolalidad sérica.
Control hemodinámico.
Las disminuciones del volumen venoso son detectadas por los receptores de distensión en las aurículas y en las grandes venas, y los
descensos de la presión arterial son detectados en los barorreceptores del cayado aórtico y el glomus carotídeo. El arco aferente son los nervios vago y
glosofaríngeo hacia el núcleo solitario de la médula oblonga (centro vasomotor). El arco eferente, desde el centro vasomotor hasta los núcleos del hipotálamo,
induce la liberación de AVP. A niveles bajos (
<
5pg/ml), la estimulación de los receptores V
2
por la AVP puede causar una retención inadecuada de agua.
A niveles elevados (
>
20pg/ml), la estimulación de los receptores V
1a
por la AVP causa vasoconstricción y participa en el reflejo barorreceptor.
(De Koeppen BM,
Stanton BA: Regulation of body fluid osmolality: regulation of water balance.
En
Koeppen BM, Stanton BA [eds.]:
Renal Physiology,
4.
a
ed. Filadelfia, Mosby
Elsevier, 2007, págs. 71-90.)