








 267 / 2894
267 / 2894

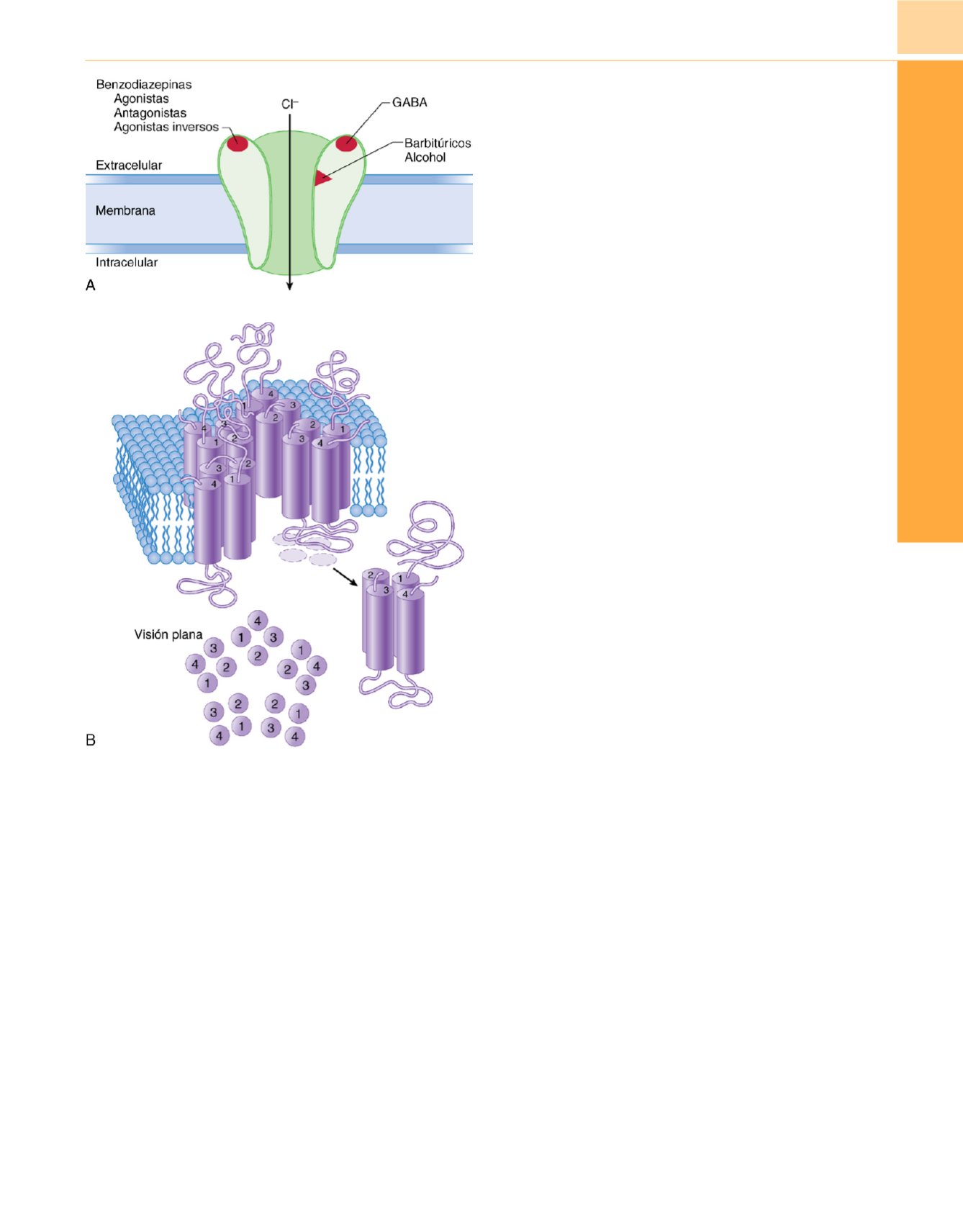
su receptor dentro del complejo canal iónico de sensible a ligando
GABA
A
y provoca un flujo de iones cloro hacia el interior de la
célula. Esta acción produce una hiperpolarización del potencial de
membrana, una característica clave de la neurotransmisión inhibi-
dora. Los fármacos que se unen a otros sitios del receptor GABA
A
facilitan la acción del GABA endógeno (v.
fig. 9-31). La mayoría de
los hipnóticos (benzodiazepinas, barbitúricos, propofol, etomidato
y probablemente la acción hipnótica de los anestésicos inhalados)
actúan mediante potenciación del GABA endógeno en el canal
iónico sensible a ligando GABA
A
.
Bombas iónicas
Otro tipo de proteína excitable de membrana es la bomba iónica.
La bomba sodio-potasio adenosina trifosfatasa (ATPasa) es quizá
la bomba iónica más conocida para el anestesiólogo porque se
inhibe con digital. El líquido extracelular tiene una concentración
alta de sodio y baja de potasio, mientras que el líquido intracelular
tiene una concentración alta de potasio y baja de sodio. Debido a
que en reposo el nervio es selectivamente permeable al potasio
pero no al sodio, el potasio se desplaza al espacio extracelular
creando una carga extracelular neta positiva y una carga intracelu-
lar neta negativa. Los potenciales de acción activan los canales de
sodio, permitiendo al sodio desplazarse al espacio intracelular a
favor de un gradiente químico y eléctrico combinado. A continua-
ción, la bomba sodio-potasio ATPasa bombea rápidamente sodio
fuera de la célula, intercambiándolo por potasio hasta que la célula
alcanza su composición catiónica y su gradiente eléctrico origina-
les. Los fármacos que actúan en las bombas iónicas alteran la pro-
porción de catión intracelular/extracelular, alterando el potencial
de reposo de la membrana.
La digital actúa inhibiendo la bomba sodio-potasio ATPasa.
Esto tiene especial importancia en la célula miocárdica, donde el
intercambio sodio-potasio-ATPasa es sustituido por un intercam-
bio sodio-calcio más lento, aumentando la concentración intrace-
lular de calcio. La función miocárdica de bombeo mejora porque
el calcio aumenta la contractilidad miocárdica.
Segundos mensajeros
La unión de un fármaco a un receptor no produce sus efectos clí-
nicos instantáneamente. Más bien hay una serie de rápidos fenó-
menos bioquímicos que relacionan la unión al receptor con los
efectos clínicos definitivos. Estos fenómenos bioquímicos se deno-
minan segundos mensajeros. Es importante que el anestesiólogo
conozca los principios generales de acción del segundo mensajero,
porque las alteraciones en el acoplamiento a segundos mensajeros
pueden alterar la efectividad de un fármaco.
Numerosos receptores de membrana se acoplan a su segundo
mensajero mediante proteínas G, que son moléculas reguladoras
intermediarias. El acoplamiento de las proteínas G a complejos
hormona-receptor requiere energía en forma de trifosfato de gua-
nosina (GTP). La hidrólisis de GTP asociado a proteína G a difos-
fato de guanosina (GDP) está regulada por otro conjunto de
proteínas denominadas
proteínas RGS
. Después de la interacción
del receptor con la proteína G, se activa la reacción bioquímica en
la cascada efectora. Existen proteínas G estimuladoras (p. ej., G
s
,
G
q
) e inhibidoras (p. ej., G
i
, G
o
). El efecto fisiológico está determi-
nado por la proteína G específica y por la consiguiente respuesta
celular. Las proteínas G son heterotriméricas, formadas por tres
subunidades:
a
,
b
, y
g
. Las interacciones entre el receptor y la
proteína G producen la disociación de las subunidades de proteína
G en
a
y
bg
. La subunidad
a
de la mayoría de las proteínas G
confiere especificidad entre receptor y efectores. Aunque original-
mente se creía que las subunidades
bg
eran simplemente proteínas
G de anclaje a la membrana celular, ahora está claro que las subu-
nidades
bg
disociadas son capaces de estimular directamente
segundos mensajeros. Además, las subunidades
bg
participan en
el anclaje de cinasas reguladoras a la membrana celular, provo-
cando un aumento de la fosforilación de los receptores de
membrana.
Uno de los sistemas de segundo mensajero mejor conoci-
dos es el sistema adenilciclasa. En este sistema, los complejos
Principios básicos de farmacología
267
9
Sección II
Farmacología y anestesia
© ELSEVIER. Fotocopiar sin autorización es un delito
Figura 9-31
Sección transversal del complejo receptor benzodiazepina-ácido
g
-aminobutírico (GABA) (que es el prototipo de complejo canal iónico sensible
a ligando). La unión de los agonistas benzodiazepinas al canal iónico sensible a
ligando GABA
A
facilita la acción del GABA endógeno. Esto aumenta el flujo de
ión cloro inhibidor en el sistema nervioso central.
A
, Dibujo esquemático del
complejo canal iónico sensible a ligando GABA
A
en el que se observan los
sitios del receptor para benzodiazepinas y GABA, así como los distintos
lugares del receptor para barbitúricos y alcohol.
B
, Modelo del receptor
GABA
A
y complejo proteína canal iónico de cloro formado por un
heterooligómero con cinco subunidades,
a
,
b
,
g
, así como polipéptidos
d
o
ρ
.
Cada subunidad tiene cuatro supuestos dominios que cruzan la membrana
(numerados de 1 a 4, representados por
cilindros
). (
A
, de Berkowitz DE:
Cellular signal transduction.
En
Scwinn DA [ed.]:
Scientific Principles of
Anesthesia,
vol. 2. Filadelfia, Current Medicine 1998;
B
, de Firestone L,
Quinlan J, Gyulai F: Mechanisms of anesthetic action.
En
Scwinn DA [ed.]:
Scientific Principles of Anesthesia,
vol. 2. Filadelfia, Current Medicine 1998.
)