








 297 / 2894
297 / 2894

nápticas excitadoras mediadas por receptores glutamato (v. cap. 1
) 219.
Los pocos experimentos sobre los efectos de los anestésicos volá-
tiles en la PLP han obtenido resultados en cierto modo contradicto-
rios: el halotano, el enflurano y el isoflurano no bloquean la
inducción PLP in vivo, la ketamina s
í 217. Por el contrario, el isoflu-
rano bloquea la PLP en el corte de hipocampo potenciando la
inhibición mediada por receptor GABA
A
( fig. 10-13 ) 220 .La DLP, un
debilitamiento dependiente del uso de las conexiones excitadoras
que es efectivamente una contrapartida homeostática de la PLP,
también es bloqueada por el isofluran
o 220 .La contradicción entre
estos hallazgos in vivo e in vitro podría reflejar diferencias en el
equilibrio entre circuitos excitadores e inhibidores activados por
el estímulo eléctrico empleado. El etomidato, a concentraciones
amnésicas, bloquea la PLP aumentando los subtipos específicos de
receptores GABA
A 119mientras que la ketamina bloquea la PLP en
animales anestesiados con uretano mediante bloqueo de los recep-
tores NMD
A 221,222.
Circuitos con actividad espontánea
La actividad neuronal espontánea disminuye por diversos anesté-
sicos volátiles tanto in vivo como en cortes de la corteza cerebral.
Este efecto es muy dependiente del receptor GABA
A
y pronun-
ciado incluso con concentraciones «sedantes» baja
s 77. Estos resul-
tados indican que los anestésicos volátiles pueden causar algunos
efectos (p. ej., sedación) mediante acción cortical directa porque
los cortes cultivados carecen de aferencias subcorticales. No obs-
tante, es posible que los cambios en las tasas brutas de activación
neuronal no sean un indicador cuantitativo preciso de función
cognitiva superior. También se han estudiado los efectos anestési-
cos en los circuitos implicados en la locomoción, un generador de
patrón central bien estudiado. Los resultados con isoflurano en
la preparación de médula espinal de lamprea indican que la médula
espinal es la diana principal de la inmovilidad provocada por anes-
tésico volátil
223 .Ritmos y simulaciones
El encéfalo genera de modo constante ritmos eléctricos complejos
con una frecuencia entre fracciones y cientos de hertzios. Aun
que no conocemos bien sus funciones fisiológicas, el concepto
predominante es que reflejan, sirven o constituyen un procesa-
miento de orden superior fundamental en el encéfalo de mamí-
fero. Su modulación por anestésicos merece atención. Los ritmos
g
y los ritmos
u
se han investigado in vitro e in vivo en el contexto
de los mecanismos anestésicos.
Ritmos
g
El isoflurano baja la frecuencia de las oscilaciones
g
provocadas
(30-80Hz, también denominados ritmos de «40 Hz») en el ser
human
o 224 .Estudios de las oscilaciones
g
in vitro sugieren que su
frecuencia depende de la constante de tiempo de desintegración de
las corrientes sinápticas mediadas por receptor GABA
A
en redes
inhibidora
s 225 .El isoflurano ralentiza los ritmos
g
en cortes de
hipocamp
o 226y de cerebro neocortica
l 227en un grado similar al del
ser human
o 224 ,lo que hace sospechar un vínculo entre efectos
a nivel de circuito y de receptor. No obstante, es improbable que la
interacción entre anestésicos y efectos en redes conductualmente
relevantes sea unitaria, porque las oscilaciones
g
provocadas con
destellos en la corteza visual primaria no cambian por los anesté-
sicos inhalatorio
s 54 ,mientras que la transferencia retrógrada de
información en frecuencias
g
entre las cortezas visual y frontal
se alter
a 228 .Ritmos
u
Los ritmos
u
, presentes en distintas estructuras corticales, son más
prominentes en el hipocampo, donde señalizan su «estado acti-
vado». Están relacionados con funciones sensitivomotoras y de
memoria en animales despiertos activo
s 229 .Un componente del
ritmo
u
(tipo I o resistente a atropina) puede verse afectado por
concentraciones amnésicas de isoflurano, así como por el no inmo-
vilizante F
6 56, lo que indica un probable efecto distintivo a nivel de
red de la amnesia inducida por anestésico. El ritmo
u
tipo II (sen-
sible a atropina) puede provocarse bajo anestesia y el halotano lo
ralentiza y potenci
a 230 .Los ritmos
g
y
u
están interconectados en
vías complejas aunque no está nada clara la naturaleza de su modu-
lación por los anestésicos ni tampoco su trascendencia.
Modelos y simulaciones
A escala atómica, el modelado de la interacción de las moléculas
anestésicas con proteínas tipo diana ha definido «motivos de
unión» que caracterizan las cavidades de unión anfífilas para las mo
Anestésicos inhalatorios: mecanismos de acción
297
10
Sección II
Farmacología y anestesia
© ELSEVIER. Fotocopiar sin autorización es un delito
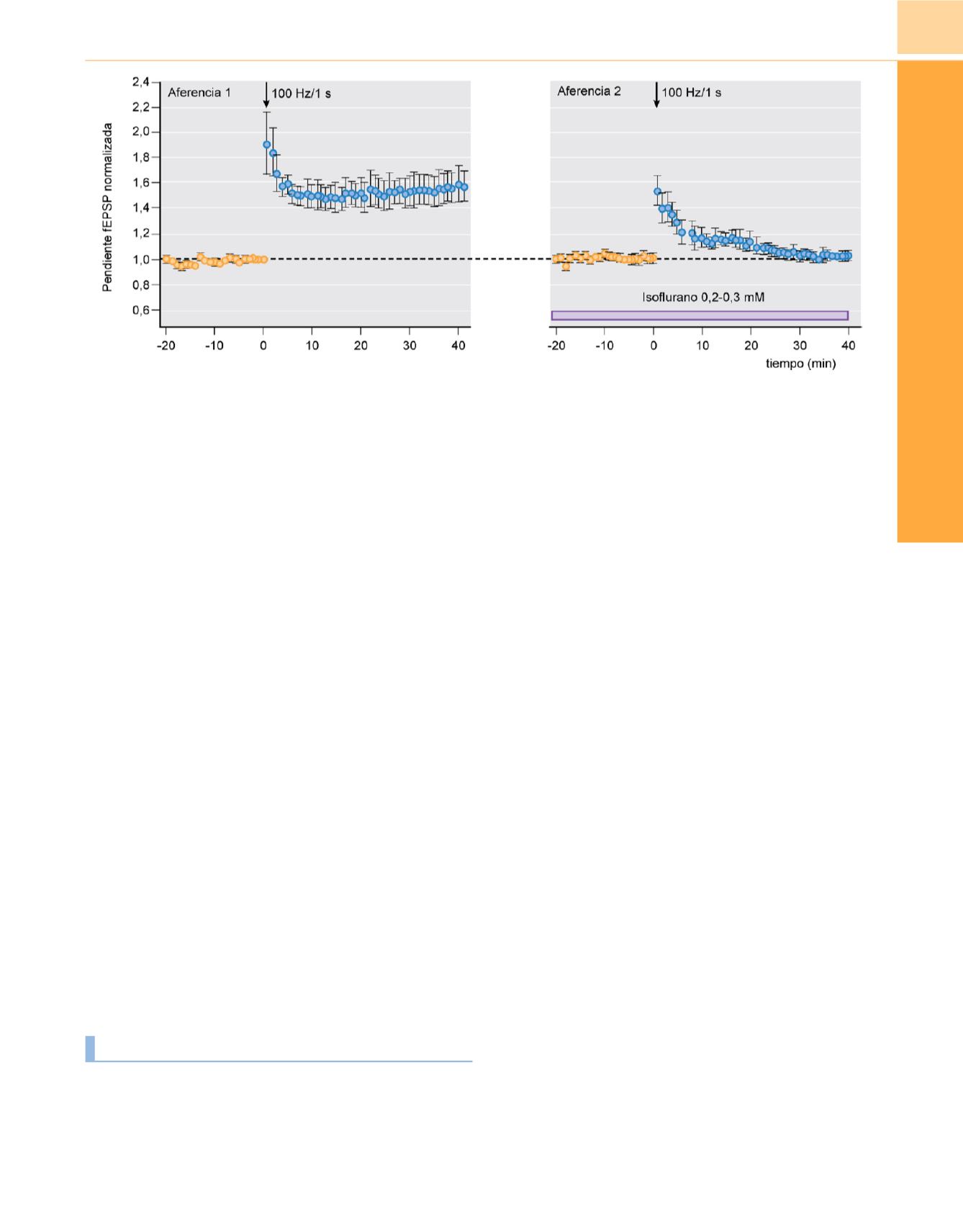
Figura 10-13
El isoflurano bloquea la inducción de plasticidad sináptica (modelo in vitro de aprendizaje y memoria). La potenciación a largo plazo (aumento de
la potencia sináptica) en cortes de hipocampo provocada por estimulación tetánica de sinapsis excitadoras se bloquea con 300
m
M de isoflurano.
(Reproducida
con autorización de Simon W y cols.: Isofluorane blocas synaptic plasticity in the Mouse hippocampus.
Anesthesiology
94:1058-1065, 2001.)