








 45 / 2894
45 / 2894

El sistema nervioso autónomo
45
2
Sección I
Fisiología y anestesia
© ELSEVIER. Fotocopiar sin autorización es un delito
intracelulares, lo que altera la actividad celular. En la aurícula del
corazón, la activación de receptores muscarínicos induce libera-
ción de potasio e hiperpolarización de la membrana celular. Esta
última hace que la conducción sea más lenta, atenuando o anu-
lando el efecto de los marcapasos auriculares. En las glándulas, la
entrada de calcio o sodio, o de ambos, incrementa la actividad
intracelular y la secreción por parte de la célula. De forma similar,
su entrada en células de músculo liso hace que éste se contraiga.
Los receptores muscarínicos se encuentran en neuronas cen-
trales y periféricas. Una misma célula puede presentar receptores
muscarínicos con efectos tanto de excitación como de inhibición.
Los autorreceptores presinápticos se conocen mejor en el sistema
simpático que en el parasimpático. Los receptores muscarínicos
presinápticos inhiben la liberación de acetilcolina por parte de las
neuronas parasimpáticas posganglionares, mientras que los recep-
tores nicotínicos presinápticos aumentan su producción.
Dada la complejidad del acoplamiento, la respuesta del
sistema muscarínico es lenta. De hecho, tras la aplicación de ace-
tilcolina no se registra respuesta alguna durante segundos o
minutos. El efecto sobrevive no obstante a la presencia del agonista.
Incluso aunque el transmisor se destruya rápidamente, la secuencia
de episodios que inicia induce una respuesta celular que dura
varios minutos. Los receptores muscarínicos son desensibilizados
por fosforilación dependiente del agonista, en un mecanismo
similar al descrito para los receptores
b
-adrenérgicos.
Neurotransmisión no adrenérgica
no colinérgica (NANC) en el sistema
nervioso autónomo
Componentes NANC, tales como monoaminas, purinas, aminoá-
cidos y polipéptidos, forman parte del SNA. Otros posibles trans-
misores detectados en nervios perivasculares, mediante técnicas
histoquímicas e inmunohistoquímicas, son ATP, adenosina, VIP,
sustancia P, 5-hidroxitriptamina (5-HT), neuropéptido Y (NPY) y
péptido relacionado con el gen de la calcitonina (CGRP, por sus
siglas inglesas). En estudios inmunocitoquímicos se ha demostrado
que en un mismo nervio se puede localizar más de un transmisor,
efectivo o potencial. Las combinaciones más comunes de transmi-
sores en los nervios perivasculares son noradrenalina, ATP y NPY
en nervios simpáticos
( fig. 2-14 ), acetilcolina y VIP en nervios para-
simpáticos
( fig. 2-15 )y sustancia P, CGRP y ATP en los nervios
sensoriales-motores. Muchos de estos transmisores actúan en
cotransmisión, lo que implica síntesis, almacenamiento y libera-
ción de más de un transmisor por parte de un nervio. Inicialmente,
la diversidad de transmisores liberados en distintas combinaciones
parece aleatoria y en cierto modo desconcertante. Sin embargo,
poco a poco se ha ido perfilando un patrón de ordenación que
clarifica la situación. Los nervios autónomos presentan un código
químico, en virtud del cual las neuronas con una determinada
función contienen diferentes combinaciones de transmisores
71 .La cotransmisión y la neuromodulación se han aceptado
como mecanismos de control de los nervios autónomos. Para
determinar qué transmisores actúan simultáneamente en los mis
mos nervios, es necesario constatar que, al ser liberada, cada sus-
tancia actúa postsinápticamente sobre su receptor específico para
generar una respuesta.
En numerosos nervios simpáticos perivasculares se ha con-
firmado que la noradrenalina y el ATP actúan como cotransmiso-
res y son liberados por los mismos nervios, aunque actúan sobre
a
1
-adrenorreceptores y P
2
-purinorreceptores, respectivamente,
para inducir vasoconstricción (v.
fig. 2-14 ) 72,73. El ATP, del que antes
se creía que sólo actuaba como amortiguador eléctrico de la
noradrenalina cargada, actualmente se considera mediador de la
contracción por medio de los receptores P
2
a través de los canales
del calcio dependientes del voltaj
e 74. El componente rápido de la
contracción parece ser mediado por estos purinorreceptores, mien-
tras que la noradrenalina mantiene la contracción del músculo
actuando sobre el receptor adrenérgico
a
1
mediante los canales del
calcio activados por el receptor. El ATP se almacena en vesículas
de las varicosidades nerviosas. Es liberado a la hendidura sináptica
por exocitosis y, a continuación, se une a los receptores purinérgi-
cos postsinápticos. Después se decompone, para formar adenosina
por acción de las ATPasas y las 5’-nucleotidasas de membrana. La
adenosina es posteriormente captada en la neurona presináptica,
donde el ATP es resintetizado e incorporado a las vesículas para su
ulterior liberació
n 75.
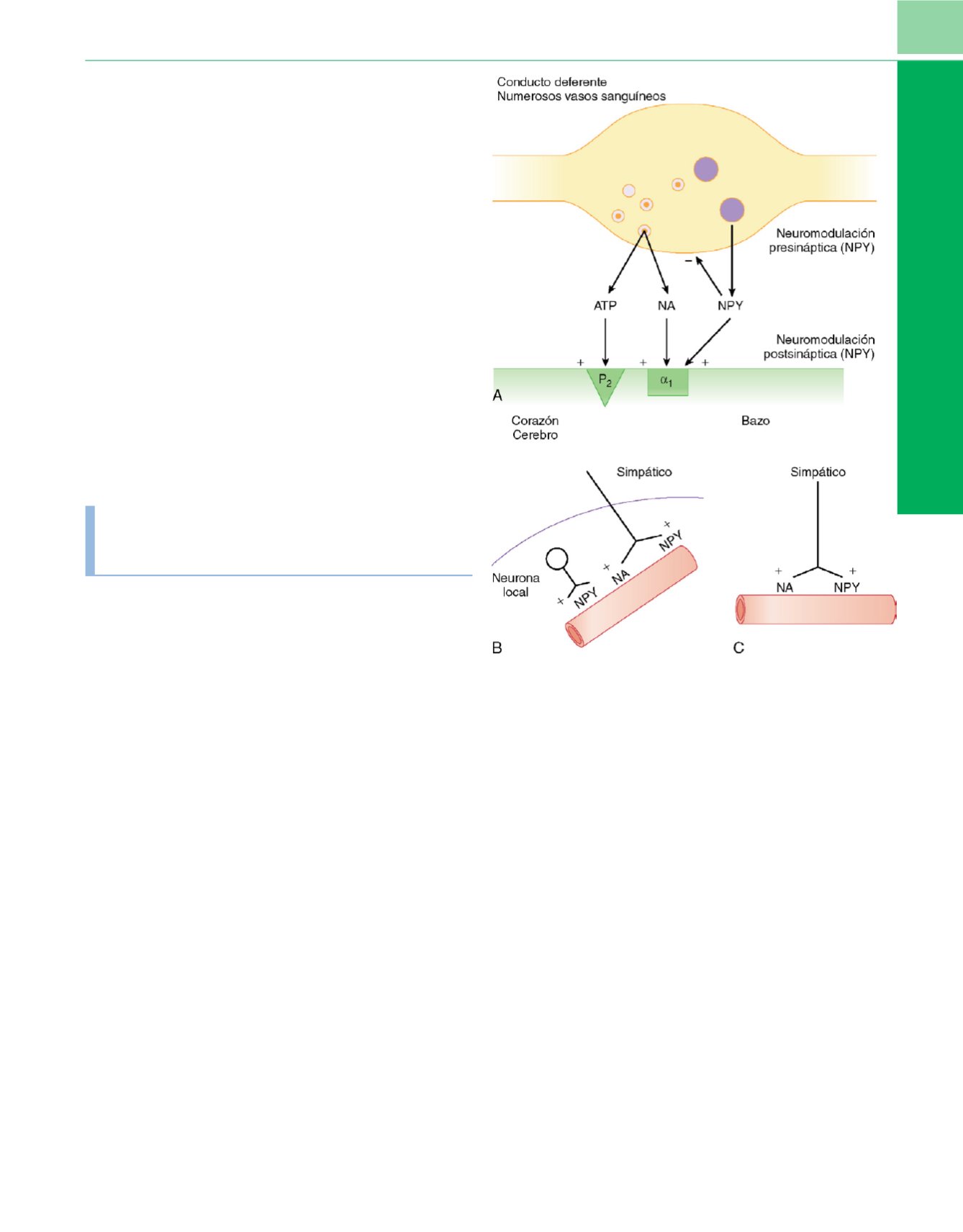
Figura 2-14
Representación esquemática de diferentes interacciones que se
producen entre el neuropétido Y (NPY) y el adenosintrifosfato (ATP), y la
noradrenalina (NA) liberada de cada varicosidad de un nervio simpático.
A,
El
diagrama muestra lo que sucede en el conducto deferente y en numerosos
vasos sanguíneos, donde la NA y el ATP, probablemente liberados por
pequeñas vesículas granulares, actúan de forma sinérgica para contraer (+) el
músculo liso por medio de los receptores adrenérgicos
a
1
y los
P
2
-purinorreceptores, respectivamente.
B
y
C,
Neurotransmisión simpática en
el corazón y el cerebro
(B)
y en el bazo
(C)
. (
De Lincoln J, Burnstock G:
Neuralendothelial interactions in control of local blood flow.
En
Warren J (ed.)
:
The Endothelium: An Introduction to Current Research.
Nueva York, Willey-
Liss, 1990, pág. 21
.)