








 685 / 2894
685 / 2894

Anestésicos locales
685
20
Sección II
Farmacología y anestesia
© ELSEVIER. Fotocopiar sin autorización es un delito
(Kelvin), F es la constante de Faraday, [K
+
] es la concentración de
potasio intracelular (i) y extracelular (e). Por tanto, para el potasio:
E
k
= −58 log30, o−85,7 mV
Para el sodio la situación es la contraria, ya que su concentración extra-
celular es más elevada y su potencial de Nernst (E
Na
) es de alrededor de
+60mV. Durante el potencial de acción, la membrana de la célula ner-
viosa modifica transitoriamente su mayor permeabilidad del K
+
al Na
+
,
por lo que el potencial de membrana cambia de negativo a positivo,
para volverse negativo de nuev
o 8 .La evolución de este cambio de poten-
cial y los sucesos que lo producen se muestran en la
figura 20-6 ,que
proporciona las bases necesarias para comprender cómo los anestésicos
locales bloquean la conducción nerviosa.
Los iones atraviesan las membranas por medio de unas pro-
teínas especiales, denominadas canales iónico
s 9. La conformación
de estos canales a menudo es sensible al potencial de membrana, de
modo que en las membranas de las células nerviosas, tanto los
canales del Na
+
como los del K
+
son activados hacia una conforma-
ción «abierta» durante la despolarización de la membrana. Además,
los canales del sodio se cierran (conformación «inactivada») tras su
activación inicial. Una pequeña despolarización de la membrana que
se extienda a lo largo de un axón desde una zona de membrana
excitada, por ejemplo, comenzará a abrir tanto los canales de Na
+
como los de K
+
. Sin embargo, los canales de Na
+
se abren con más
rapidez y los iones de Na
+
que entran en el nervio lo despolarizan
aún más (v.
fig. 20-6 ), originando la abertura de más canales de Na
+
e incrementando más la corriente interna de Na
+
( fig. 20-7 ). Esta
secuencia de hechos continúa durante la
fase despolarizante
, hasta
que se desactivan algunos de los canales de Na
+
y se abren los sufi-
cientes canales de potasio para cambiar el balance de la corriente,
que resulta en una corriente neta de salida que produce la
repolari-
zación
de la membrana (v.
fig. 20-7 ). La modificación de las concen-
traciones de Na
+
y K
+
tras un potencial de acción es mínima en las
fibras mielinizadas de gran calibre, pero es de hasta el 10% en los
axones pequeños no mielinizados. Los iones de Na
+
y de K
+
que
entran y salen de la célula, respectivamente, como resultado de este
proceso, son repuestos mediante la bomba de Na
+
/K
+
.
Las despolarizaciones que son demasiado débiles como para
activar un número suficiente de canales de Na
+
que produzcan una
corriente neta hacia el interior se encuentran por debajo del
umbral
de excitabilidad de lamembrana.El valor exacto del umbral varía entre
las diferentes regiones de la célula y también varía con el tiempo. Por
ejemplo, inmediatamente después de un potencial, cuando todavía
algunos de los canales de Na
+
se encuentran inactivados y algunos
canales de K
+
aún están activados, el umbral se encuentra por encima
del valor de «reposo» y la membrana es «refractaria» a la estimulación.
Sin embargo, con el paso del tiempo los valores originales del umbral
de reposo se restablecen al cesar la inactivación de los canales de Na
+
y al retomar los canales de K
+
su conformación cerrada.
El potencial de acción es una onda de despolarización que
se propaga a lo largo del axón mediante el continuo acoplamiento
entre regiones excitadas y regiones no excitadas de la membrana.
La corriente iónica (la corriente de acción) que penetra en el axón
en la región excitada despolarizada atraviesa el axoplasma y aban-
dona el axón a través de la membrana circundante, despolarizando
pasivamente la región adyacente (v.
fig. 20-3).Aunque esta
corriente
de circuito local
se extiende en ambas direcciones a partir de la zona
excitada, la región posterior al impulso que acaba de ser despola-
rizada se vuelve completamente refractaria, por lo que la propaga-
ción del impulso es unidireccional.
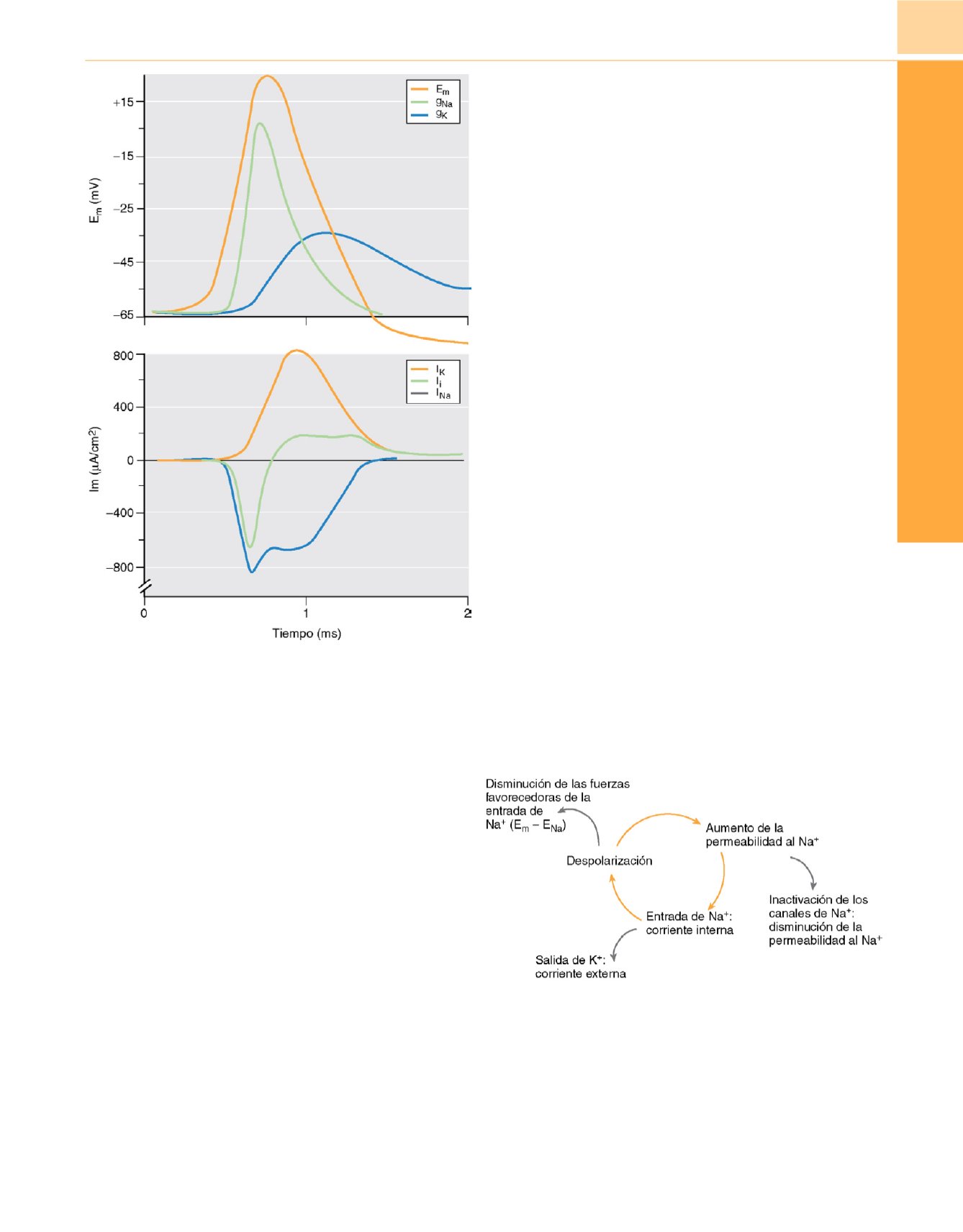
Figura 20-6
Potencial de membrana (E
m
) y conductancia iónica del sodio
(g
Na
) y del potasio (g
K
) dependientes del voltaje, que causan las
correspondientes corrientes en la membrana (I
Na
e I
K
) durante la propagación
de un potencial de acción. Estas relaciones, obtenidas a partir de los estudios
originales de Hodgkin y Huxley en el axón del calamar gigante (v. Hodgki
n 8 )son válidas para las fibras nerviosas de casi todos los vertebrados e
invertebrados. La corriente iónica total (I
i
), que es la suma de I
Na
y de I
K
, se
dirige hacia el interior (valores negativos) durante la fase despolarizante del
potencial de acción y hacia el exterior (valores positivos) durante la fase de
repolarización.
Figura 20-7
El potencial de acción puede entenderse en términos de las
relaciones cíclicas entre los factores que contribuyen a la fase despolarizante,
la fase regenerativa y la fase repolarizante pasiva. Los factores
positivos
(flechas amarillas)
aumentan la tasa de despolarización por medio de un
«sistema de retroalimentación positiva», de modo que cada elemento del
ciclo estimula al siguiente. Los factores
negativos (flechas grises)
reducen
la tasa de despolarización al disminuir u oponerse a los factores positivos,
por lo que, como resultado, la salida de K
+
domina finalmente el flujo iónico
y repolariza la membrana.