








 285 / 2894
285 / 2894

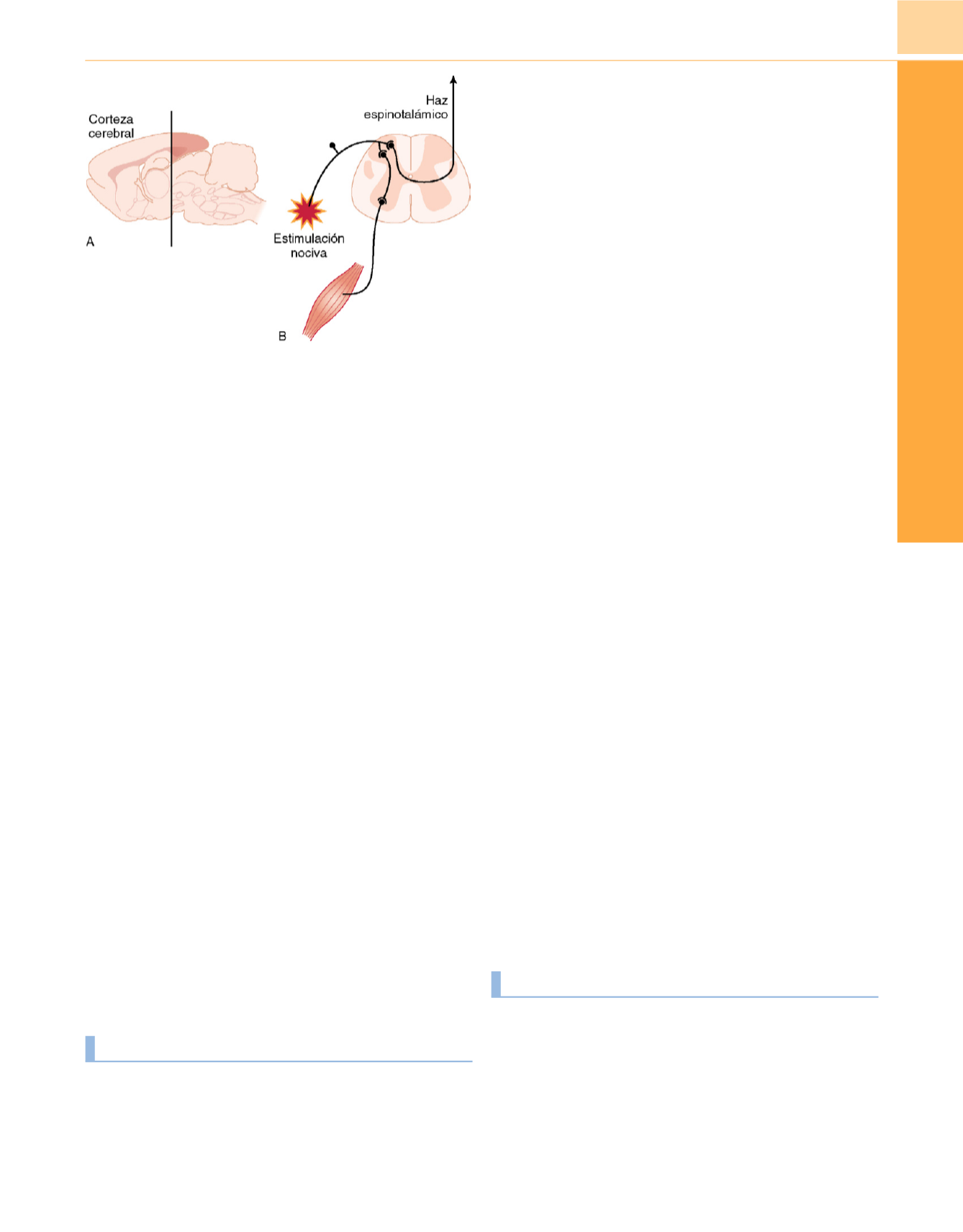
La separación quirúrgica del prosencéfalo de la médula espinal
en la rata confirmó que la inmovilización implica la supresión prin-
cipalmente del arco reflejo de retirada defensivo a nivel de la médula
espinal
( fig. 10-4 ) 18 .Estudios recientes se han centrado en los mecanismos por los
que los anestésicos inhalatorios producen inmovilidad. Un método
farmacológico para identificar las contribuciones a nivel de recepto-
res a la CAM consiste en la administración intratecal de antagonistas
selectivos de los receptores neurotransmisores in vivo. Este método
reveló que las acciones en los receptores glutamato de tipo
N
-metil-d-
aspartato (NMDA
) 25y en los receptores glicin
a 26contribuyen a la
inmovilización provocada por isoflurano, mientras que las acciones
en los receptores GABA
A
pueden ser irrelevante
s 27 .Es probable que
los receptores glicina y GABA
A
tengan escasa implicación en la
inmovilización por ciclopropan
o 26, lo que se debe al menos en parte
al antagonismo de los receptores NMD
A 28. Otros métodos de inves-
tigación han descartado otras dianas posibles. La sensibilidad a fár-
macos no inmovilizadores descartó la participación de la inhibición
de los receptores de acetilcolina nicotínicos en la inmovilizació
n 29 ,y los ratones transgénicos resistentes a anestésicos confirmaron que
los receptores GABA
A
con subunidades
a
1
y
a
3
no contribuyen a la
acción inmovilizante del isofluran
o 30,31. Por el contrario, los ratones
mutantes sin canales K
2P
TASK-1, TASK-3 y TREK-1 tienen una
CAM más alta para los anestésicos volátiles, pero no para los anes-
tésicos intravenoso
s 32-34, lo que indica la participación de estos
canales, probablemente por un mecanismo presináptic
o 35 .A pesar
de que se ha identificado el punto anatómico macroscópico para la
inmovilidad y se ha implicado a dianas moleculares específicas, sigue
sin haber una hipótesis integral del mecanismo de la inmovilidad
provocada por anestésicos inhalatorios (v. ref.
36para revisión).
Inconsciencia
La consciencia, como cualidad de la mente, es fácil de entender pero
difícil de definir (v. cap. 29). Como proceso neurobiológico reali-
zado en el encéfalo, está al alcance de la ciencia objetiva pero aún
se conoce poco. La pérdida de la consciencia (o hipnosis) es un
hecho distintivo del inicio de la anestesia, que ofrece la posibili-
dad excitante de que los fármacos anestésicos puedan ser útiles
para comprender la consciencia mediante los «correlatos nervio-
sos de la consciencia
» 37 .Aunque no hay una definición científica
universalmente aceptada, la consciencia consiste en estados sub-
jetivos, cualitativos e internos de «noción o alerta
» 38o de «alerta
explícita
» 39 .El conocimiento del mecanismo por el que los anestésicos
anulan la consciencia está surcado por teorías de la generación de
la consciencia, muchas sin sustento experimental ni predicciones
comprobables. Por el contrario, los planteamientos neurocientíficos
de la consciencia pueden analizarse de modo experimental. La
«teoría talámica» de la anestesia plantea que el mecanismo de la
inconsciencia es una desaferenciación somatosensitiva por acción
anestésica en el tálamo
40 .En apoyo de esta hipótesis, el isoflurano
hiperpolariza y cortocircuita las neuronas talámica
s 41 ,una acción
coherente con la alteración de la transferencia talámica de infor-
mación observada in viv
o 42 .La imagen funcional del encéfalo
humano muestra una supresión preferente de la actividad talámica
por algunos, anestésicos (aunque no todos), y esto ha dado lugar
a la hipótesis del «cambio talámico
» 43 .No obstante, la pérdida de
consciencia se produce con un intervalo muy estrecho de concen-
traciones anestésica
s 44,45, habitualmente inferiores a 0,5 CA
M 46,47,
mientras que los efectos cuantificables en el tálamo aparecen por
encima de este intervalo de concentración y son, por lo general,
progresivos (tipo dímero) y no bruscos (tipo cambio). Una teoría
integral de la inconsciencia provocada por anestésicos debe ser más
incluyente que un bloque simple de transferencia de información
a través del tálamo para resultar coherente con la evidencia dispo-
nibl
e 48y también debe explicar la supresión de la actividad cortical
endógena generada sin estimulación externa.
La neurociencia contemporánea ha reemplazado la visión
cartesiana de una estructura encefálica diferenciada como el centro
de la consciencia por el concepto de que la consciencia requiere la
integración de la información entre múltiples regiones encefálicas
a través de redes cerebrales de gran escal
a 49,50. La interferencia
anestésica con la sincronía y coherencia operativa de estas redes
con la consiguiente alteración de la conectividad funcional cortical,
como ocurre en el sueño natural
51y en estados vegetativos, podría
provocar «desunión cognitiva
» 52con pérdida de conscienci
a 38. La
inconscienci
a 53estaría caracterizada no por la ausencia, sino por
la desintegración del procesamiento cortica
l 39 .Aunque el mecanismo
de «unión» (formación de la unidad de percepción) es dudoso, la
sincronía de la actividad neuronal en el intervalo de 40 a 90Hz
a través de áreas corticales conectadas funcionalmente (habitual-
mente denominada ritmo
g
o de 40Hz) es un candidato viable. Hay
información en animales
54-56y en el ser human
o 44que implica a la
actividad en la banda
g
a lo largo de la corteza como diana a nivel
de red de los anestésicos generales. Las acciones anestésicas en
el procesamiento de la información cortical consisten probable-
mente no sólo en suprimir las respuestas, sino en reducir la com-
plejidad y variabilidad reflejada de modo paradójico en un aumento
de la fiabilidad y precisión de las respuestas provocada
s 57.
Aprendizaje y memoria
La amnesia anterógrada, uno de los efectos anestésicos favorables,
se consigue conmenores concentraciones anestésicas (
∼
0,25 CAM)
que las necesarias para la inconsciencia (
∼
0,5 CAM). Quizá el
análogo más próximo en roedores para explicar la memoria en el ser
humano es el aprendizaje espacial dependiente del hipocampo.
Puede evaluarse con distintos modelos experimentales como el
condicionamiento del miedo al contexto
( fig. 10-5). Otros modelos
Anestésicos inhalatorios: mecanismos de acción
285
10
Sección II
Farmacología y anestesia
© ELSEVIER. Fotocopiar sin autorización es un delito
Figura 10-4
Los anestésicos inhalatorios producen inmovilidad a nivel
medular.
A,
La descerebración mediante extirpación del prosencéfalo por
delante de la
línea negra gruesa
no altera la CAM de isoflurano en ratas,
lo que indica que la inmovilización por anestésico volátil no depende de la
corteza cerebral.
B,
Los anestésicos suprimen la respuesta refleja de retirada
defensiva a la estimulación nociva transmitida al asta dorsal por nervios
sensitivos a nivel medular. Los estudios actuales se centran en identificar los
sustratos moleculares, celulares y anatómicos de este efecto.
(
A
modificada
de Rampil IJ, Masson P, Singh H: Anesthetic potency [MAC] is independent of
forebrain estructures in the rat.
Anesthesiology
78:707-712, 1993.)