








 286 / 2894
286 / 2894

de aprendizaje, como el condicionamiento del miedo al tono, son,
por el contrario, independientes del hipocampo. El isoflurano y el
F6 no inmovilizante inhiben el aprendizaje dependiente del hipo-
campo con aproximadamente la mitad de la concentración necesa-
ria para el aprendizaje independiente del hipocamp
o 58 .Los efectos
anestésicos en la función hipocámpica deben estar implicados en
la supresión de la memoria explícita, porque las concentraciones de
anestésicos que inhiben la memoria explícita en el ser humano
son igual de bajas que las que empeoran la memoria implícita
47 .Los efectos en otras estructuras, como la amígdala, pueden ser
cruciales para el deterioro anestésico de la memoria implícita y de
otros tipo
s 59.
La comparación de la potencia amnésica de cinco anestési-
cos inhalatorios con el modelo de evitación inhibidor reveló que el
óxido nitroso era el amnésico más potente y el halotano, el menos
potente (expresado en fracciones CAM), con los éteres halogena-
dos de potencia intermedi
a 60 .Es difícil atribuir la amnesia a meca-
nismos celulares específicos porque los anestésicos inhalatorios
actúan en múltiples dianas celulares a concentraciones amnésicas.
Tampoco está claro si la inhibición del aprendizaje y la de la
memoria con fármacos con distintas afinidades por los receptores
comparten mecanismos comunes en cierto nivel de integración. La
comparación con fármacos más selectivos aporta información útil.
Hay abundante evidencia de que los ritmos
u
(4-12Hz) son impor-
tantes para el aprendizaje y la memoria dependientes del hipo-
camp
o 61 .Las benzodiazepina
s 62y los cannabinoide
s 63ralentizan y
suprimen los ritmos
u
hipocámpicos en proporción a su capacidad
para alterar el aprendizaje dependiente del hipocampo. El isoflu-
rano y el F6 no inmovilizante tienen efectos comparables en los
ritmos
u
a concentraciones amnésicas a pesar de sus perfiles dife-
rentes a nivel de receptor y efectos opuestos en la sedació
n 56 .Por
tanto, las alteraciones en la sincronía neuronal aportan un sustrato
común a nivel de red del deterioro de la memoria. La sincronía
entre los ritmos
u
en amígdala e hipocampo que existe durante
la recuperación de la memoria del miedo indica que este principio
podría aplicarse también a otras formas de memoria y a su altera-
ción por los anestésico
s 64 .Igual que con otros componentes del
estado anestésico, quedan por descubrir los mecanismos exactos
de alteración de la memoria por los anestésicos y de la propia
memoria.
Sedación
La sedación (definida como un descenso de actividad, vigilia, lucidez
y/o vigilancia) y su continuidad conductual que es la hipnosis se
consiguen con concentraciones anestésicas bajas (
<
0,5 CAM), las
que producen amnesia. No hay una separación causal ni clínica
entre sedación e hipnosis. Por el contrario, a pesar de que es difícil
separar la sedación de la amnesia, la evidencia obtenida con anes-
tésicos intravenosos apoya la existencia de sustratos separados,
aunque solapados, para estos dos criterios de valoració
n 20,65. Los
mecanismos implicados en estos efectos conductuales son proba-
blemente similares a los de fármacos menos promiscuos, para los
que los métodos genéticos han sido informativos. Una mutación
activada de un aminoácido (H101R) en ratones que hace insensible
a la subunidad
a
1
del receptor GABA
A
a la regulación por benzo-
diazepinas confiere resistencia a los efectos amnésicos y sedantes de
286
Farmacología y anestesia
II
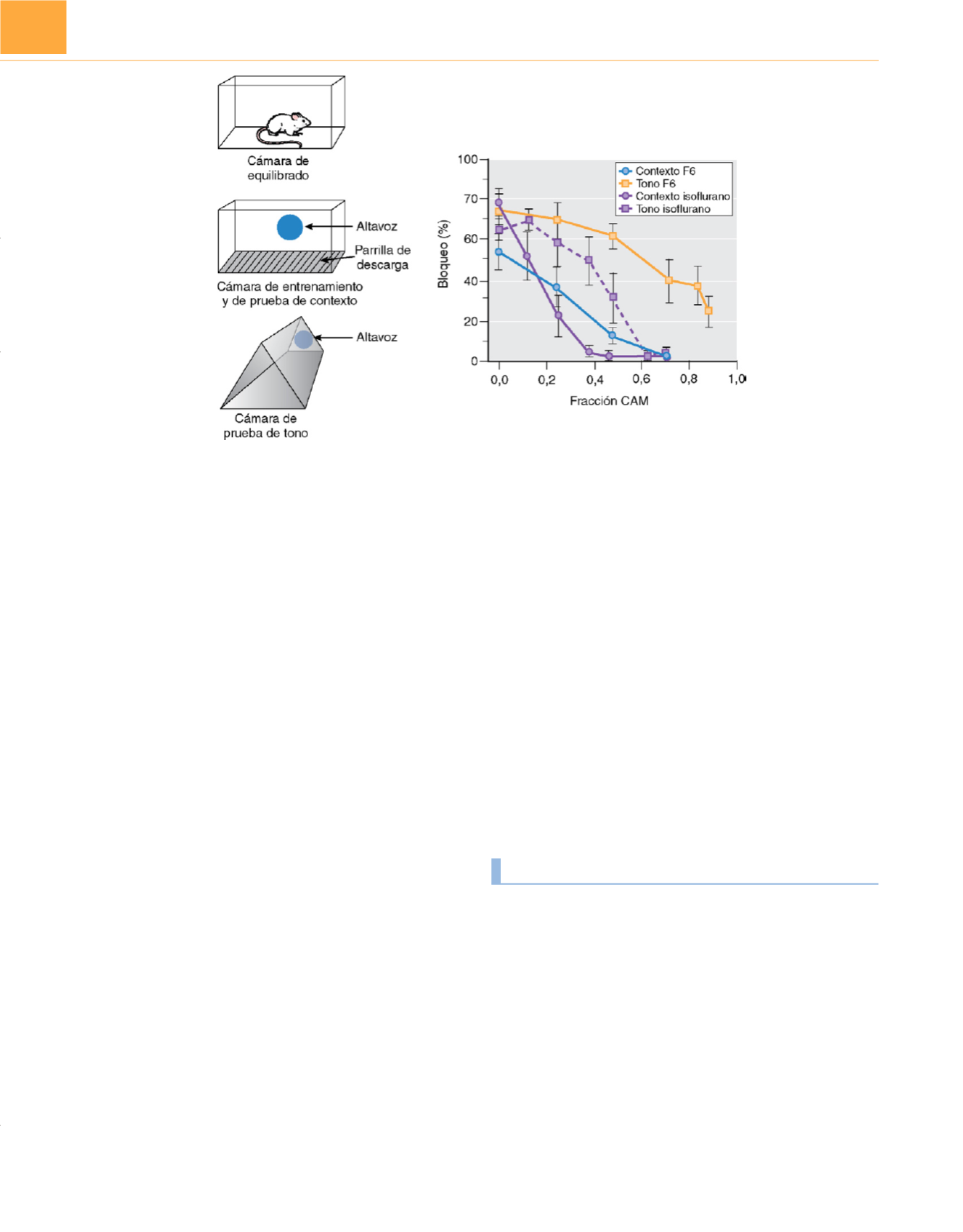
Figura 10-5
Sensibilidad diferencial de distintos tipos de aprendizaje a los anestésicos y no inmovilizantes. El bloqueo en anticipación de un estímulo nocivo
es un indicador de aprendizaje en ratas. Menos bloqueo indica menos aprendizaje.
Izquierda,
El protocolo de aprendizaje implica pre-equilibrado de las ratas
en la cámara de equilibrado a isoflurano o al no inmovilizante F6 a la concentración deseada antes de introducirlas en la cámara de entrenamiento. Para
estudiar la memoria de contexto, la cámara de entrenamiento y la cámara de prueba son idénticas. Para estudiar la memoria de tono, el entrenamiento y la
prueba tienen lugar en distintas cámaras.
Derecha,
El aprendizaje dependiente del hipocampo (condicionamiento de miedo al contexto
, símbolos cerrados
)
se inhibe por el isoflurano (
círculos violetas
) a menores concentraciones que el aprendizaje independiente del hipocampo (condicionamiento de miedo al tono,
cuadrados violetas
). Esta sensibilidad diferencial es igual con el no inmovilizante F6(
círculos azules
y
cuadrados amarilllos
para contexto y tono,
respectivamente). (
Panel izquierdo
modificado con autorización de Eger El 2
nd
y cols.: Isoflurane antagonizes the capacity of flurothyl or
1,2-dichlorohexafluorocyclobutane to impair fear conditioning to context and top tone.
Anesth Analg
96:1010-1018, 2003;
panel derecho:
los puntos de
datos reconstruidos de Dutton RC, y cols.: Short-term memory resists the depressant effect of the nonimmobilizer 1-2-dichlorofluorocyclobutane (2N) more
than long-tern memory.
Anesth Analg
94:631-639, 2002 y Dutton RC y cols.: The concentration of isofluorane required to suppress learning depends on the
type of learning.
Anesthesiology
94:514-519, 2001.
)